緒言
魚類の色覚を考慮して、漁具の色を設定することは、漁業の種類によっては、漁獲効率を高める上で極めて重要と考えられる。魚類の色覚を知るための研究手法は、行動学的手法と電気生理学的手法に大別される。
行動学的手法の先駆けとして、 Frishch ( 1925 )はGobio fluviatilis (コイ科)を用いた学習実験により、魚類における二元説を証明した。彼は黒・灰・白・赤の皿のうち、必ず赤皿で餌をやることを繰り返すことで、実験魚に餌を入れなくても赤皿に接近する行動を条件づけることに成功した。続いて白から黒にいたる約20種位の灰色の程度をいろいろ変えても必ず赤によってくることを見い出だした。もし魚が色盲ならある程度の灰色にも反応するはずであるが、必ず赤皿に反応することより魚は赤色を認めると結論された。同様の実験から、赤以外の色でも色覚を有するという結果を得ている。また彼は魚が弱光下では色の弁別ができなくなることを指摘している。この時の網膜を固定して顕鏡すると、錐体はのびて桿体が外限界膜に接近し、色を弁別しうる程度の明るさでは錐体が外限界膜に接近していることから、錐体が色彩感覚に用いられること(二元説)を証明した。田村( 1977 )によると、錐体と桿体とは機能が異なり、前者は明器 ( photocyte )ともいわれ、明るい時に働き、多くの場合色彩の弁別の能力があり、錐体を用いているときの視覚(錐体視-明るいところでの視覚)は視精度がすぐれている。これに反して桿体は暗器 ( scotocyte )ともいわれ、うす暗い時に働き、色彩の弁別はできなく、 桿体視覚(うす暗い所での視覚)は視精度が劣る。このように環境の明るさにより感光細胞の分業が行われているとする説を二元説 ( duplicity theory )という。これは広く脊椎動物一般にあてはまる説であるとされる。Hamburger ( 1926 )は魚類の色覚に補色関係があることを証明した。ヒトの目には補色というものがある。すなわち赤色光と緑色光、青色光と黄色光といったものを適当に混ぜ合わせると 白色に見える。コイ科の魚でもこれらの光の組み合わせは白色と区別できなくなり、魚の色覚に補色関係があることを学習法で証明した。川本信之( 1959 )は主として海産稚魚イシダイ、カワハギ、サワラ、 ボラ、メジナ、ウナギ、メダカ等を用いて集魚灯の基礎的研究を行った。直径1mの木製水槽の周囲を8区画とし、その上部に白色並びに7色のフィルターを用いた等照度の光を点じ、暗室においてその中央に稚魚を入れ、魚がいかなる色光の室に最も多く入るかを測定して、走光性の大小を決定した。大多数の魚類は青と緑とに最大に集まり、赤と紫とに最小であり、フィルターを用いていない60Wマツダの電球に対しては、集魚率はフィルターを通した色光よりも概して小であることを見た。さらにウナギのような夜行性の魚が赤と紫に最も多く集まり、青と緑とに最小であることを指摘している。McClearyら( 1959 )は学習法に更に客観性をもたせるために次のような研究を行った。生きたキンギョを固定して心電図をとりながら学習する。すなわちAを見せて電撃を与え、Bを見せた時は何もしない。これをくりかえすとAを見せただけで心電図に明らかな変化が認められるようになった。この方法でキンギョは赤と緑を明暗の差としてでなく、 色調の差として弁別することが証明された。小山(1978)はアユの色光選好性について実験し、発育段階による差があるものの、黄色光>黄緑色光>青色光>赤色光の順に好むとしている。伊奈ら( 1979 )はスルメイカ内蔵、コマセアミ色素および各種合成色素を適当な担体と混合し、固体にして水槽内に置き、これをマダイがついばむ行動を 観察することにより、マダイの色に対する反応性を調べた。マダイは黄色系、橙色系、赤色系およびこれら色素系を含む混合色素系に対し反応性を示し、蛍光にも反応したことから、マダイの採餌活動は吸収の最大値が430 – 500 nmの色彩物質によって誘導されると報告した。柿元( 1985 )は網漁具に使用する網地の色彩は刺網の羅網率に影響があるとしている。Neumeyer ( 1992 )はキンギョを用いた加法混色 実験を行い、キンギョの色覚が紫外線感光錐体を含む4色型であるとしている。柿元ら( 1995 )は水槽の壁面の色彩を明度、彩度、色相の心理的3属性の要素別に変化させて魚類に選択させる方法により、魚類の色彩の選択性を調べた。この方法でマダイは低い明度を好み、緑ならば鮮やかな緑を好み、色相では青色( 450 nm付近)と黄色及び赤色( 600 – 700 nm付近)に高い選択率が見られたとしている。また、 色彩環境が生育に及ぼす影響をマダイを黄色、灰色、コンクリート地の水槽で飼育し、調べた結果、選択性が高かった黄色環境下において生育が助長されている傾向が見られたとしている。
電気生理学的手法はERGによる方法とS-電位による方法に大別される。Kobayashi ( 1962 )は海水魚を含む20種の魚類について波長の異なる光源を用いてERGを記録し、またこれを指標として魚類の生態との関連を求めた。すなわち、魚類の生息水深が増加するにつれて、分光感度の最大値が短波長の方に移行する傾向が認められ、これは水深による太陽光のスペクトル分布と相関があると述べている。また、マダイのスペクトル感度曲線は470 nmで最大値をとり、550 nmと600 nmで副次的な最大値をとることから、青に敏感な要素が著しく高いとしている。また約3 lxの弱光に順応した眼から525 nmに ピークをもつ感度曲線が得られ、650 nmより長波長の光に網膜は ほとんど感じないとしている。Granitら( 1939 )は網膜中の水平細胞の発生する電位( S-電位)を記録した。Tamuraら( 1967 )はS-電位を用いて魚類の色彩感覚を調べた。田村( 1977 )はC型S電位が多い魚種では色彩感覚がすぐれていると考え、このような魚種にボラ・コイ・フナ・ウグイ・ヤリタナゴ・オイカワ・ヒガイ・ブルーギル・ スズキ・ブリ・アカエイ・カワマス・アマゴ・ビワマス・ニジマス・ ブラウンマス・マハゼ・テラピア・ブラックバス等をあげている。クロダイ・チダイ・カツオ・ヤイト・ヒラソオダ・ネコザメではC型S-電位をが発見されておらず、これらは色盲に近いと思われるとしている。Kawamuraら( 1981 )はS-電位を用いてマグロとカジキ類が色盲であることを報告し、これらの魚類は色の違いが明るさ(明度)の違いとして知覚されうると指摘した。同様にKawamura ( 1981 )は マダイ、チダイ、クロダイの三種のタイ科の魚の網膜からC型S電位を記録し、これらの魚が色覚を有すると報告した。S電位は 553 – 640 nmの間で明確な反応が得られたことからC2型とされた。川村ら( 1991 )はマダイのL型S-電位は522 nm付近にピークがあり、波長633 nmの光に対する網膜感度はかなり低かったとしている。
他の電気生理学的研究としてMacNichol ( 1964 )はキンギョの一つの神経節細胞に電極をおき、スパイクを記録している場合に、560 nmの波長の光で刺激すると興奮し(スパイク頻度が増す)、570 nmで抑制がおこる(スパイクが出なくなる)というように、神経節細胞の反応がわずかな波長の差によりいちじるしく変化することを発見した。Tomitaら( 1967 )はコイの錐体の受容器電位のスペクトル感度を調べ、3種の感度曲線を得た。最大感度を示す波長は約462 nm、 529nm、611nmとされた。
またMarks ( 1965 )は顕微分光測定法により抽出や合成などの化学的操作を行わずに網膜中の錐体外節について感光色素の吸収スペクトルλmaxを測定することに成功した。キンギョの錐体において、 λmax= 455±15 nm (青)、λmax= 530±5 nm (緑)、λmax= 625±5 nm (赤)の異なった色素を有する3種の錐体の存在を証明した。Bowmaker ( 1991 )はキンギョにλmax= 355 – 360 nmの紫外線感光錐体を含めて、単錐体の個数がどのくらいあるかを明らかにした。これらの魚類は赤、緑、青および紫外光受容細胞を含む4色錐体系により色の受容が行われるため、その色覚は4色型といわれる。
本研究では物体色に対する魚群の行動を知るための基礎的実験として、色の3属性の中から色相に着目し、行動学的手法を用いて、 キンギョCarassius auratusとマダイPagrus majorの色相弁別実験を行った。キンギョは色覚を有することは生理学的・行動学的に明らかにされているが、色相の知覚に関する研究は見かけられない。そこで物体色の赤・緑・青に対する色彩弁別実験を行った。マダイは電気生理学的には色覚が存在するとされているが、それを行動学的に示した研究は少ない。そこで赤・緑を灰色と弁別できるかを調べることで色覚の有無を調べた。まず、第1章でキンギョの報酬と罰による赤・緑・青の色相学習実験について、第2章でキンギョの色相弁別実験について、第3章でマダイの報酬による赤・緑の色相学習実験について、総合考察において研究から得られた魚類の色覚に関する知見をまとめ、魚類の色覚研究の課題について述べる。
第1章 キンギョの色彩学習実験
第1節 材料と方法
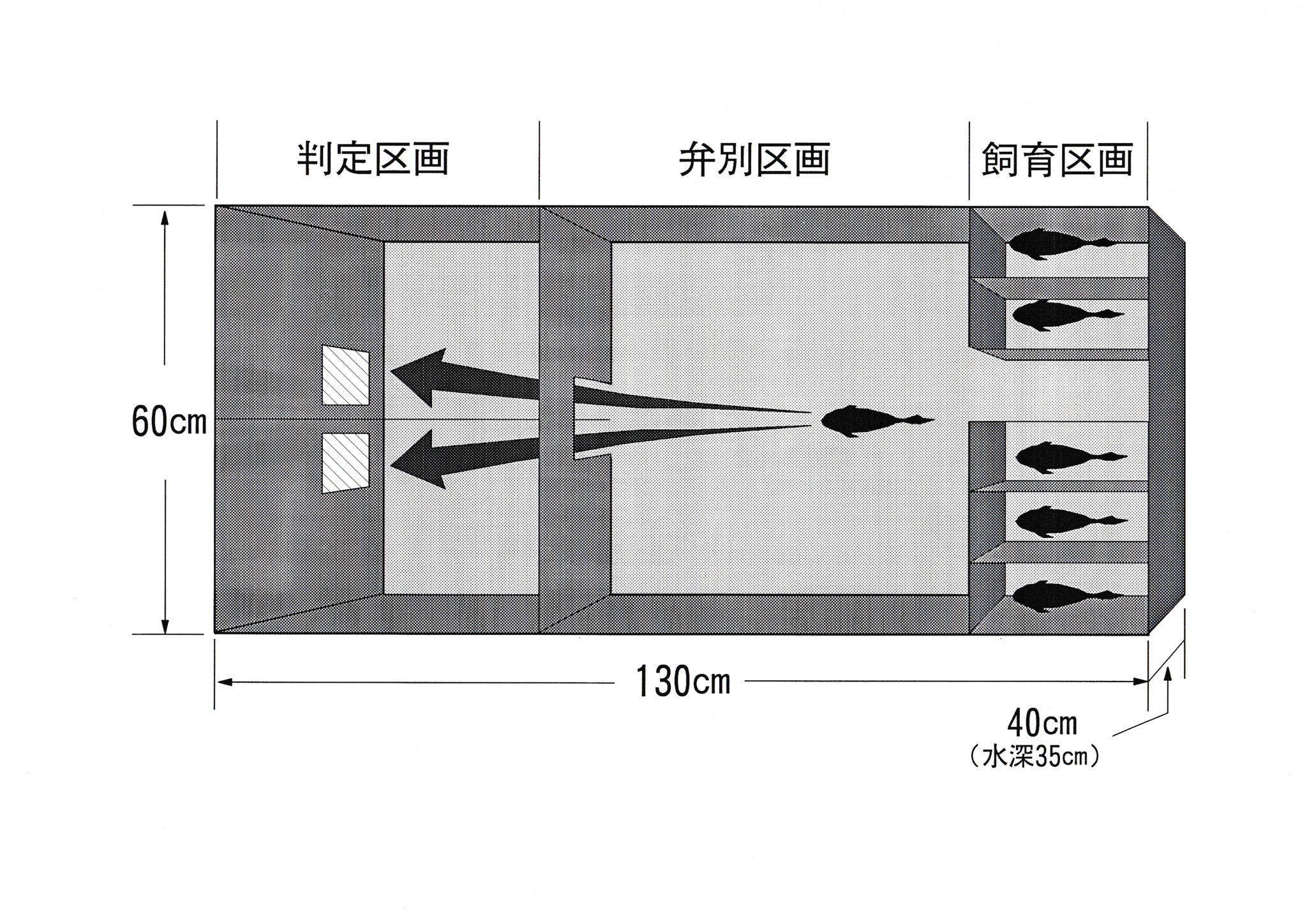
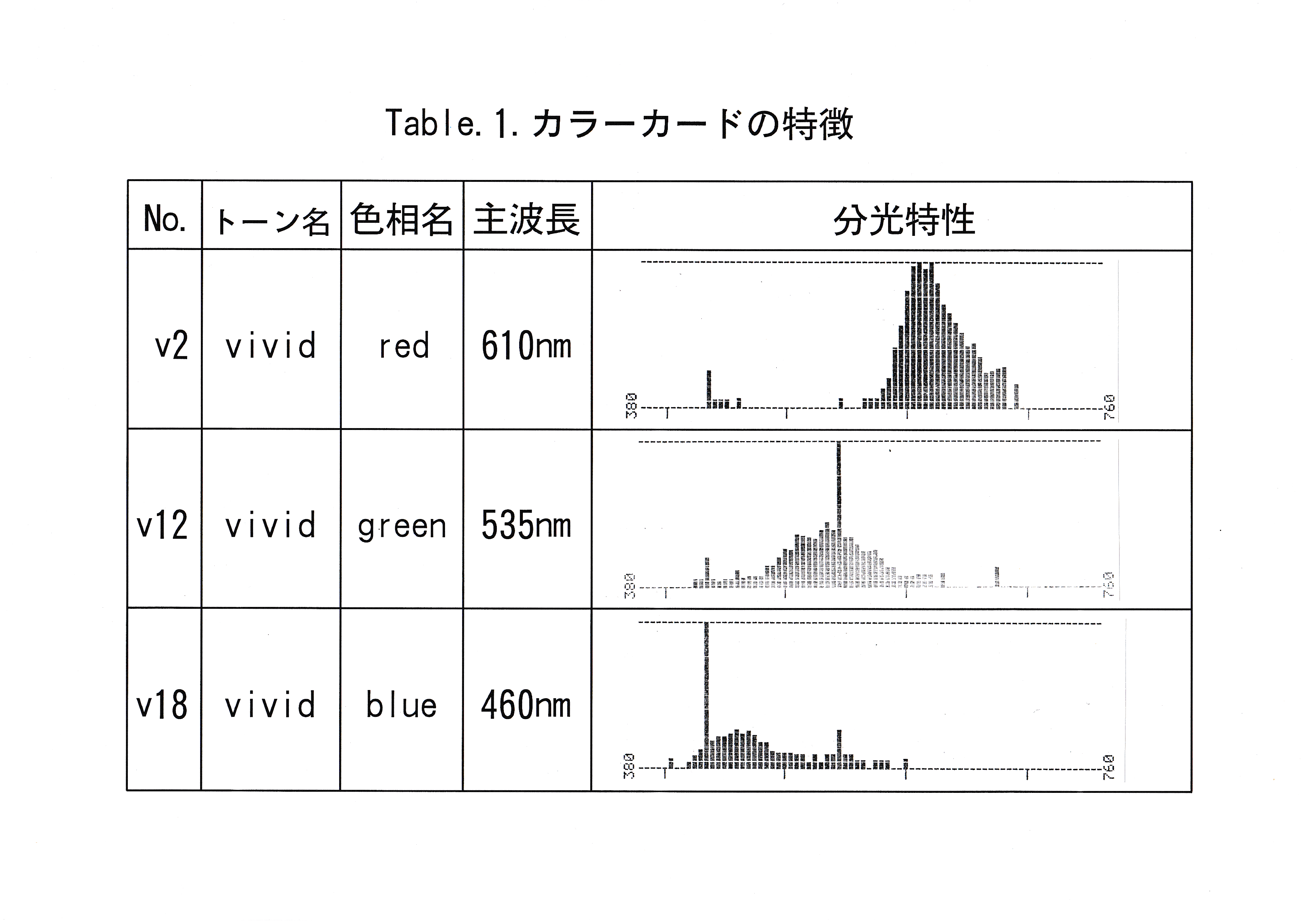
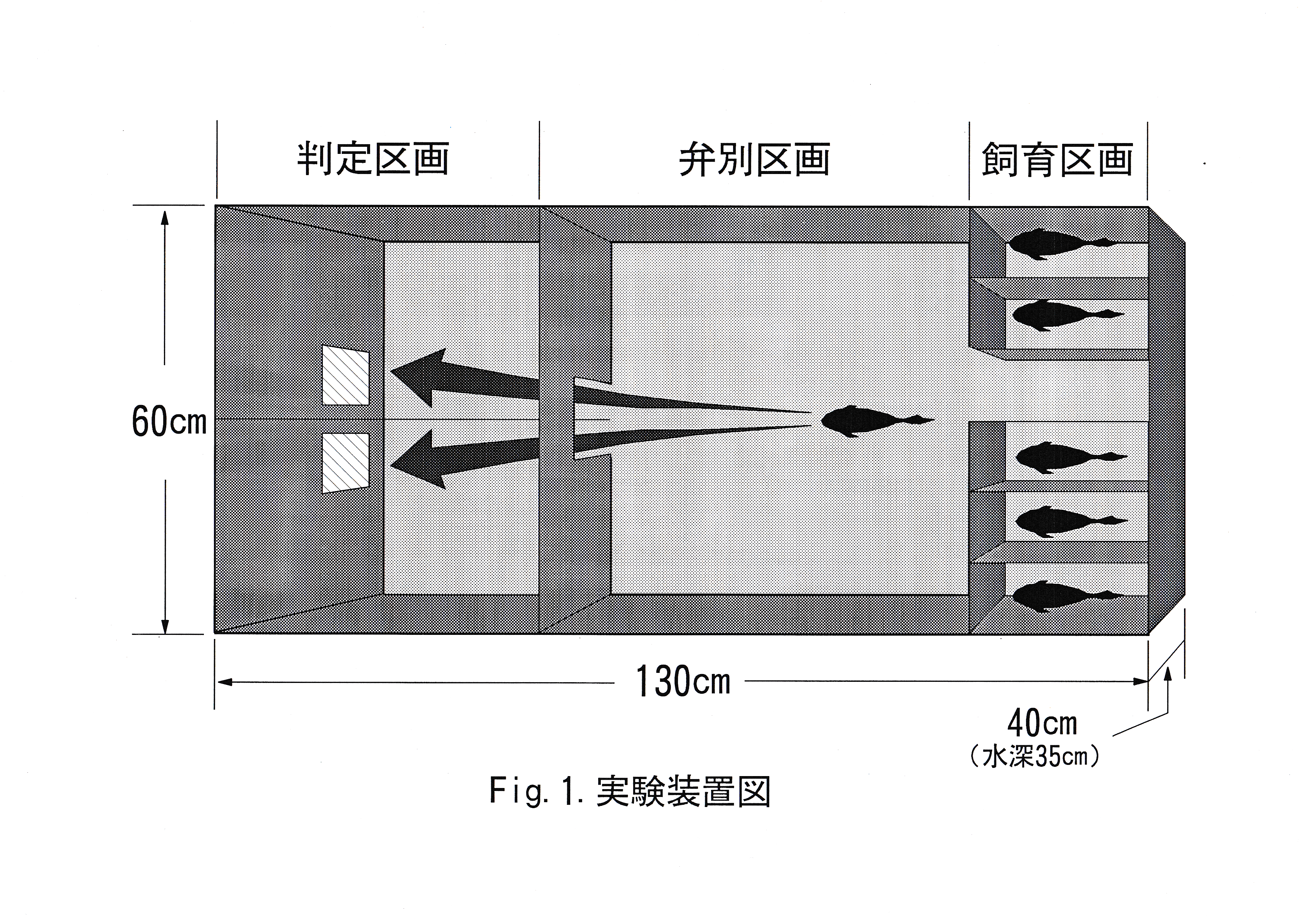
実験魚としてキンギョCarassius auratus 6尾を用いた。実験は東京水産大学、魚群行動学研究室の実験室において、平成7年4月25日から平成7年8月18日まで行った。 色の刺激としてカラーカード(日本色研事業株式会社製,新配色カード175b , v2 ・ v12 ・ v18 )を6cm四方に切り、水中に掲示できるように透明プラスチックによりコーティング(明光商会社製, L-100 mini MSパウチと、同社製100ミクロンMSパウチフィルムMP 10-90126使用)したカード3枚を用いた。用いたカラーカードの日本色研事業株式会社による色の種類( No. ,色名)をTable.1に示す。 分光放射計(東京光学機械株式会社製,分光放射計 SR-1 )を用いてパウチ後のカードの放射輝度を測定した。 実験装置図をFig.1に示す。 実験水槽は130×60×40 cmの透明 アクリル水槽を用いた。水深は35 cm一定とし、濾過装置やエア レーションにより水質・溶存酸素量を安定させた。サーモスタッド・ヒーター・クーラーを使用し、飼育時の水温を26℃一定とした。 これらの装置は濾過装置を除いて実験中は水槽内から除外した。 水槽外を黒の塩ビシートで覆い、魚に他の色覚的要素を認識させないようにした。
水槽内はアクリル板により、飼育区画・弁別区画・判定区画の大きく分けて3つの区画に仕切った。飼育区画は白のアクリル板でさらに6つの区画に分け、そこで魚を1尾ずつ飼育した。区画上面にプラスチック製無結節網地を張ることにより、実験魚が水槽の外に飛び跳ねないように配慮した。弁別区画と判定区画の間はアクリル板で仕切り、ゲート(開口部10×10cm)を設け、遠隔操作により開閉できるようにした。水槽内部は黒色に統一した。中央の仕切りにより2つの区画を設け、魚が弁別区画からゲートを通過する際に、左右の区画のどちらかにしか侵入できないようにした。侵入しようとする魚が2枚のカラーカードを同時に視認できる位置にカードを掲示板を設置した。カードを色刺激として学習しやすいようにカード掲示場所の真下に給餌口を作製した。
餌には沈降性のドライペレット(キョーリンフード工業株式会社製,ランチュウベビーゴルド,小粒)を用いた。給餌方法として、 あらかじめ餌を給餌装置の中に入れておき、実験時に遠隔操作によって給餌口につながる漏斗の中に餌を落とした。給餌は実験時にのみ行った。
実験水槽は暗幕内に置き、20 wの蛍光灯(岩崎電気株式会社製,アイライン,白色, FL20SW ) 2本を照明器具(岩崎電気株式会社製, FEC2202 )内に収容したものを照明として用いた。水槽全体を照らすために照明器具を水面中央から高さ60 cmの位置に設置した。 カード掲示位置の水中照度は159 lxであった。照明点灯後、実験魚の網膜を明順応状態とするために1時間以上順応させた後に実験を開始した。
実験開始前に飼育区画と弁別区画の間のゲートを開放し、弁別区画内に実験魚が侵入したのを確認した後にゲートを閉じ、観察者は実験結果に影響しないように暗幕の外に退去した。暗幕内の水槽上方に8mmビデオカメラを設置し、実験観察は暗幕外のモニターで行った。 実験時には弁別区画と判定区画の間のゲートを開放し、実験魚が左右2つに別れた判定区画のうち、正解のカード(正解色)が掲示してある方を選んだら正解とし、不正解のカード(不正解色)が掲示してある方を選んだら不正解とした。カードの掲示方向は左右ランダムとした。
学習のために報酬と罰を用いた。実験魚が弁別区画と判定区画の間のゲートを通過したら、ゲートを閉じる。正解した場合は報酬として餌を与え、実験魚が餌を食べ終わったらゲートを開放した。不正解の場合はただちにゲートを開放し、罰として手網で、実験魚を弁別区画に追い出した。不正解の場合は正解するまで繰り返した。
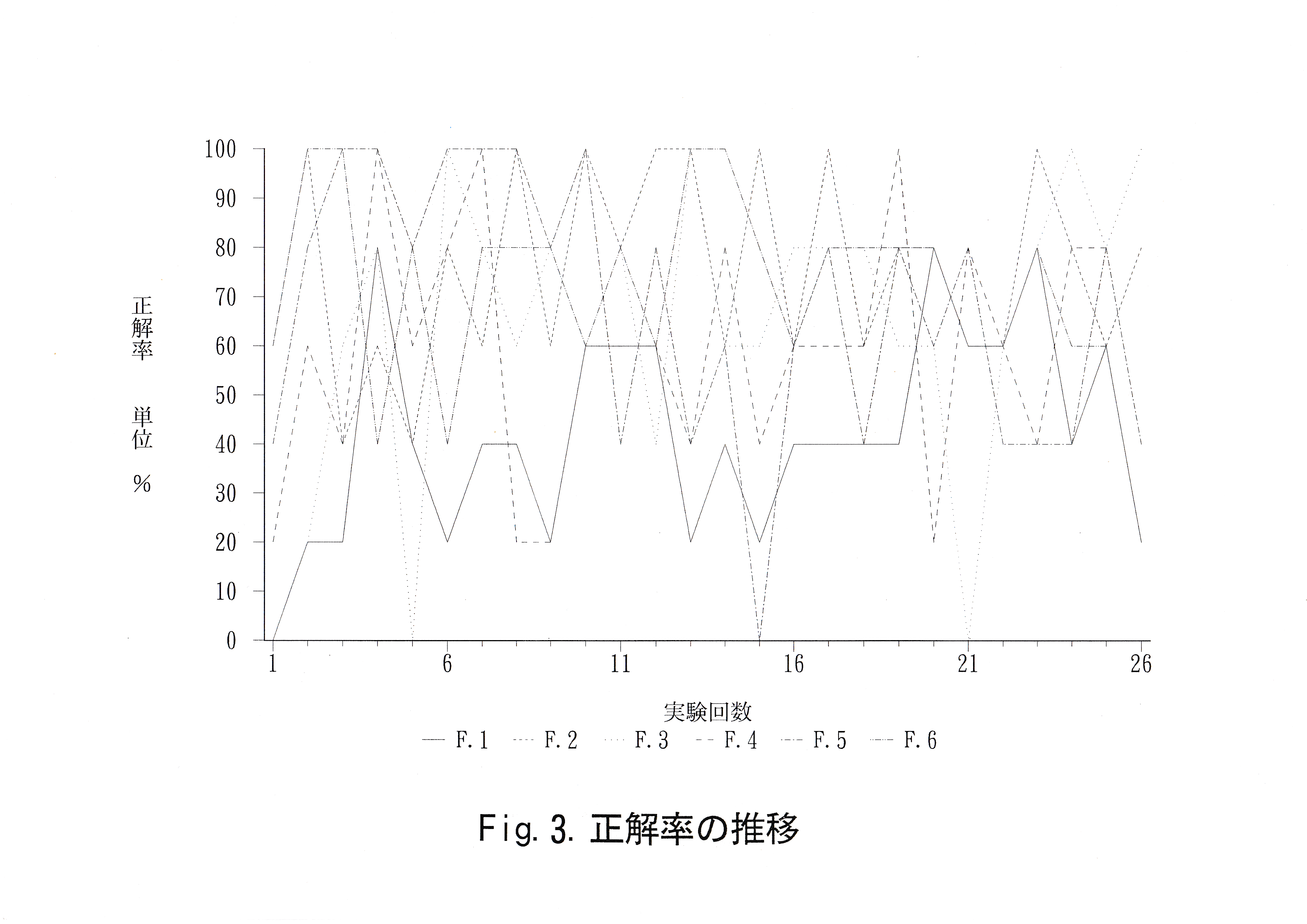
学習には赤(波長610nm ) ・緑(波長535nm ) ・青(波長460nm )の3色を使用した。これら3色のうち1色を正解色、それ以外の2色の うち1色を不正解色とする色の組み合わせは、全部で6通りある。これらの色の組み合わせ1通りにつき1尾ずつ学習を施した。 Table.2に実験魚の学習内容を示した。 学習形成の状況を把握するために実験開始時から実験魚が給餌口をつつくまでの経過時間を測定した。実験魚の学習の判定基準として、正解率を用いた。ここで正解率は実施回数に対する正解の割合と 定義した。実施回数5回を1実験とし、実験回数ごとに総経過時間・正解率の推移を示した。
第2節 結果
コーティング後のカードの放射輝度を測定した結果とカードの主波長をTable.1に示した。
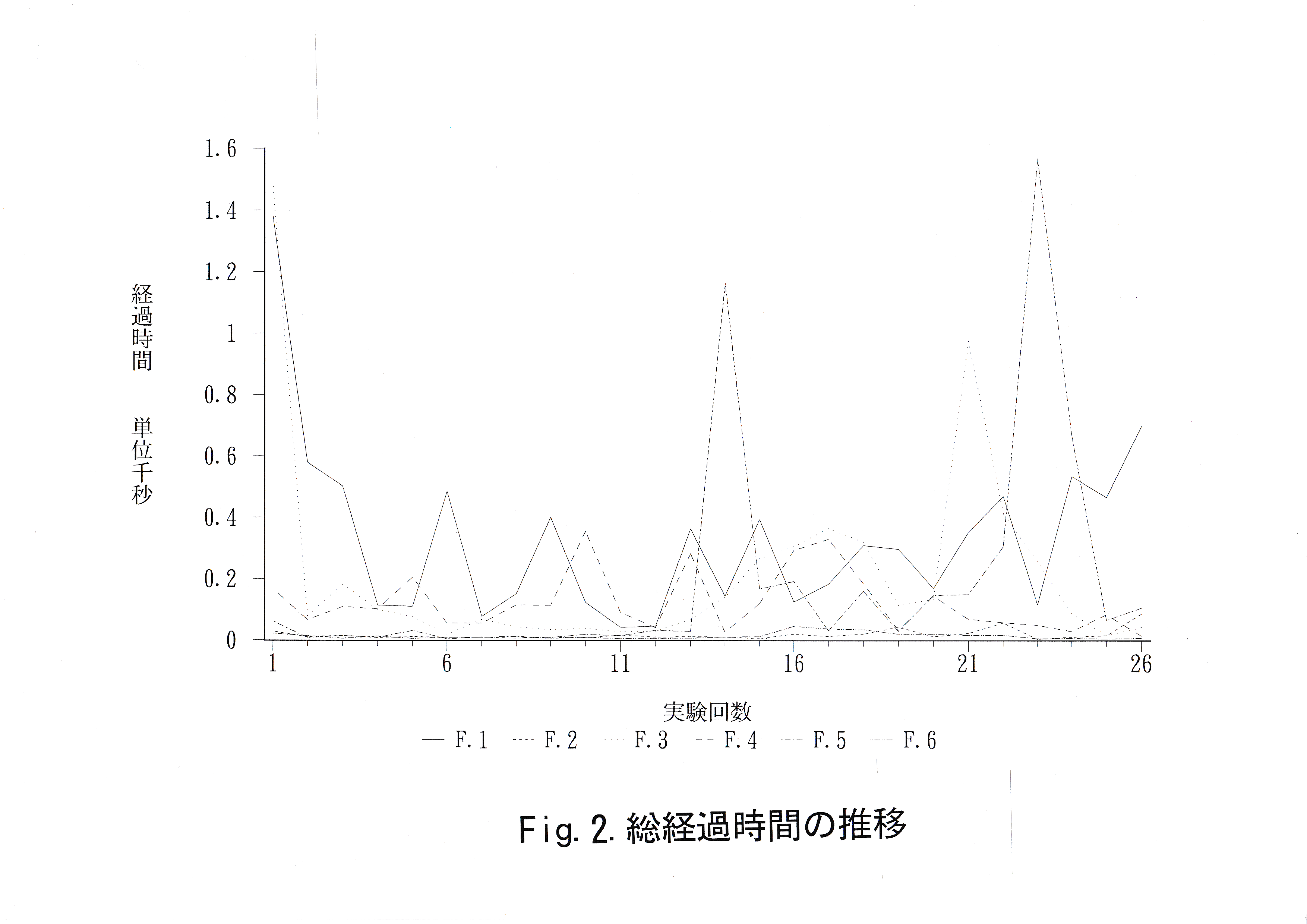
全実験魚の実験回数ごとの総経過時間の推移をFig.2に示す。 経過時間の減少は見られなかった。実験魚No . 5は実験13回目から多くの時間を要している。実験21回以降に実験魚No. 1 ・ No. 3 ・ No. 5に経過時間の増加が見られた。
実験回数ごとに正解率の推移をFig.3に示す。 実験魚No. 1は初めは赤を避ける傾向にあったが実験19回目・ 22回目に正解率80 %をとっている。実験魚No. 2は正解率60 – 100 %をとっている。実験魚No. 3は正解率に激しい変動が見られたが実験回数にして5回、 正解率100 %をとっている。実験魚No. 4は実験回数にして3回、 正解率100 %をとっている。実験魚No. 5は実験13回目以降まで 高い正解率をとっていた。実験13回目以降は実験19回目・ 21回目・ 25回目に正解率80 %をとっている。実験魚No. 6は実験回数にして4回正解率100 %をとっている。
なお、実験は段階的に難易度を上げながら行った。4月25日に実験魚を水槽内に収容してから1週間は色刺激のない状態で全区画を開放し、給餌口から餌を与えることで水槽内の地形と給餌口を覚えさせた。その後飼育区画に1尾ずつ収容し、正解色・不正解色を掲示し、弁別区画を開放し、正解の方向の給餌口に餌を落とした状態で 1尾ずつ飼育区画を開放し、給餌させた。実験魚が給餌口に餌がない状態で、ゲートの開閉や給餌装置の動作に驚かずに判定区画に進入し、給餌口をつついてから給餌できるようになったのは6月12日で、 実験結果として示したのはそれ以降の結果である。6月12日以降の変更点は、実験21回目以降に実験魚が判定区画内に進入したら ゲートを閉じたことである。さらに実験13回目からは実験魚No. 5が実験12回目修了後に死亡してしまったため、別のキンギョを実験魚として用いた結果となっている。
考察
結果からは経過時間に減少傾向が見られないが、実験開始以前には給餌口に餌をおいた状態での摂餌に1時間以上を要したことや装置の動作に驚いて判定区画に進入しようとしなかったことを考えると経過時間はかなり少なくなっていると考えられる。学習実験21回以降、実験魚No. 1 ・ No. 3 ・ No. 5に見られた経過時間の増加は、判定区画進入後にゲートが閉じることに驚いた結果と考えられ、特に実験魚No. 5は時間がかかったことから実験環境になれていない魚はより憶病になると思われる。これらの結果からキンギョは実験環境に慣れることに最も時間を要し、実験の経過時間に影響するのは音や振動などの刺激が主な原因と考えられる。
正解率は高い値をとることがあったが、安定することはなく、全体的に上昇するというよりは50 %に集束する傾向があったと考えられる。結果からは学習が形成されていないと考えられる。
井上( 1978 )はキンギョの場合、10回程度の試行で色による条件付けが可能と述べている。本実験で色の条件付けができなかった要因として以下の点が考えられる。①実験の難易度が高い。②色刺激が音や振動と比べて弱い刺激であり、正刺激として成立しにくい。③不正解でもすぐに餌が得られてしまうから、不正解が正刺激となってしまった。④色の変化を感じないため、色の学習だということがわからない。これらのなかで④については弁別実験を行うことにより確認できることから、弁別実験を行うことにした。
第2章 キンギョを用いた色相弁別実験
第1節 材料と方法
弁別実験は学習実験修了後、平成7年8月22日から平成7年8月 26日まで継続して行った。実験装置・環境は変えず、学習実験で色の学習を施したキンギョ6尾をそのまま実験魚として用いた。 弁別実験では色の刺激としてカラーカード(日本色研事業株式会社製,新配色カード175b, v2・ v3・ v4・ v5・ v6・ v7・ v8・ v9・ v10・ v11・ v12・ v13・ v14・ v15・ v16・ v17・ v18 )を6 cm四方に切り、水中に掲示できるように透明 プラスチックでコーティング(明光商会社製, L-100 mini MSパウチと、同社製100ミクロンMSパウチフィルムMP 10-90126使用)したカード17枚を用いた。用いたカラーカードの日本色研事業株式会社による色の種類( No. ,色名)をTable.3に示す。 コーティング後のカードの放射輝度を分光放射計(東京光学機械 株式会社製,分光放射計 SR-1 )を用いて測定した。この結果をもとに、正解色には学習実験で用いた色を使用し、不正解の色として正解色から不正解色までの波長域に主波長が存在するカードを用いた。これらの色を比較色と呼ぶことにする。
実験魚が学習した正解色を消去しないように、報酬のみを与えた。実験魚が弁別区画と判定区画の間のゲートを通過したら、ただちに 給餌口につながる漏斗の中に餌を落とした。実験魚は正解した場合にはすぐに餌を与えられ、不正解の場合には判定区画の反対側に進入して摂餌するという困難を課せられた。実験魚が最初に進入した区画により、正解・不正解の判定を行った。
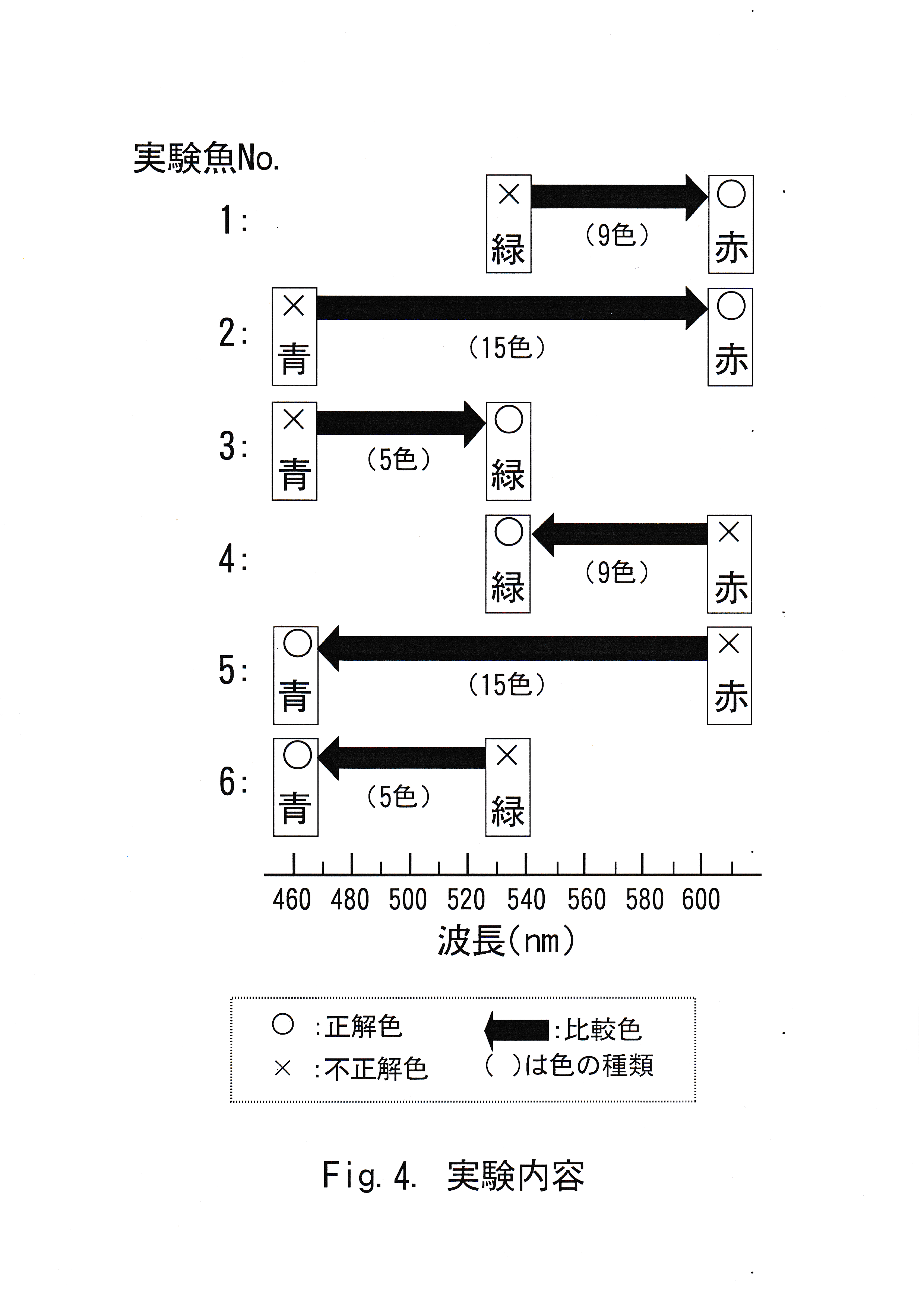
学習実験と同様に赤( v2 ) ・緑( v12 ) ・青( v18 )の3色のうち1色を正解色、それ以外の2色のうち1色を不正解色とする色の組み合わせは、全部で6通りある。これらの色の組み合わせ1通りにつき 1尾ずつ異なる実験を行った。
実験魚と学習に用いた色の組み合わせとして、 実験魚No. 1には正解色として赤( v2 )を用い、比較色として不正解色の緑( v12 )から黄色がかった赤( v3 )までの10色を用い、弁別させた。実験魚No. 2には正解色として赤( v2 )を用い、比較色として不正解色の青( v18 )から黄色がかった赤( v3 )までの16色を用い、弁別させた。実験魚 No. 3には正解色として緑( v12 )を用い、比較色として不正解色の青( v18 )から青みをおびた緑( v13 )までの6色を用い、弁別させた。 実験魚No. 4には正解色として緑( v12 )を用い、比較色として不正解色の赤( v2 )から黄色がかった緑( v11 )までの10色を用い、弁別させた。実験魚No. 5には正解色として青( v18 )を用い、比較色として 不正解色の赤( v2 )から青( v17 )までの16色を用い、弁別させた。 実験魚No. 6には正解色として青( v18 )を用い、比較色として不正解色の緑( v12 )から青( v17 )までの6色を用い、弁別させた。Fig.4に実験魚の実験内容を示した。以上の実験を1日1回ずつ5日間実施した。
弁別能力の判定には正解率を用いた。正解率は実験回数に対する正解の割合と定義した。なお実験結果では正解色と弁別させた色の波長ごとに正解率を示すため、比較色のうち主波長が等しいものについてはそれらの正解率の平均値を示した。
第2節 結果
分光放射計を用いてカードの放射輝度を測定した結果(Table.1参照)から主波長と考えられる波長をTable.3に示した。
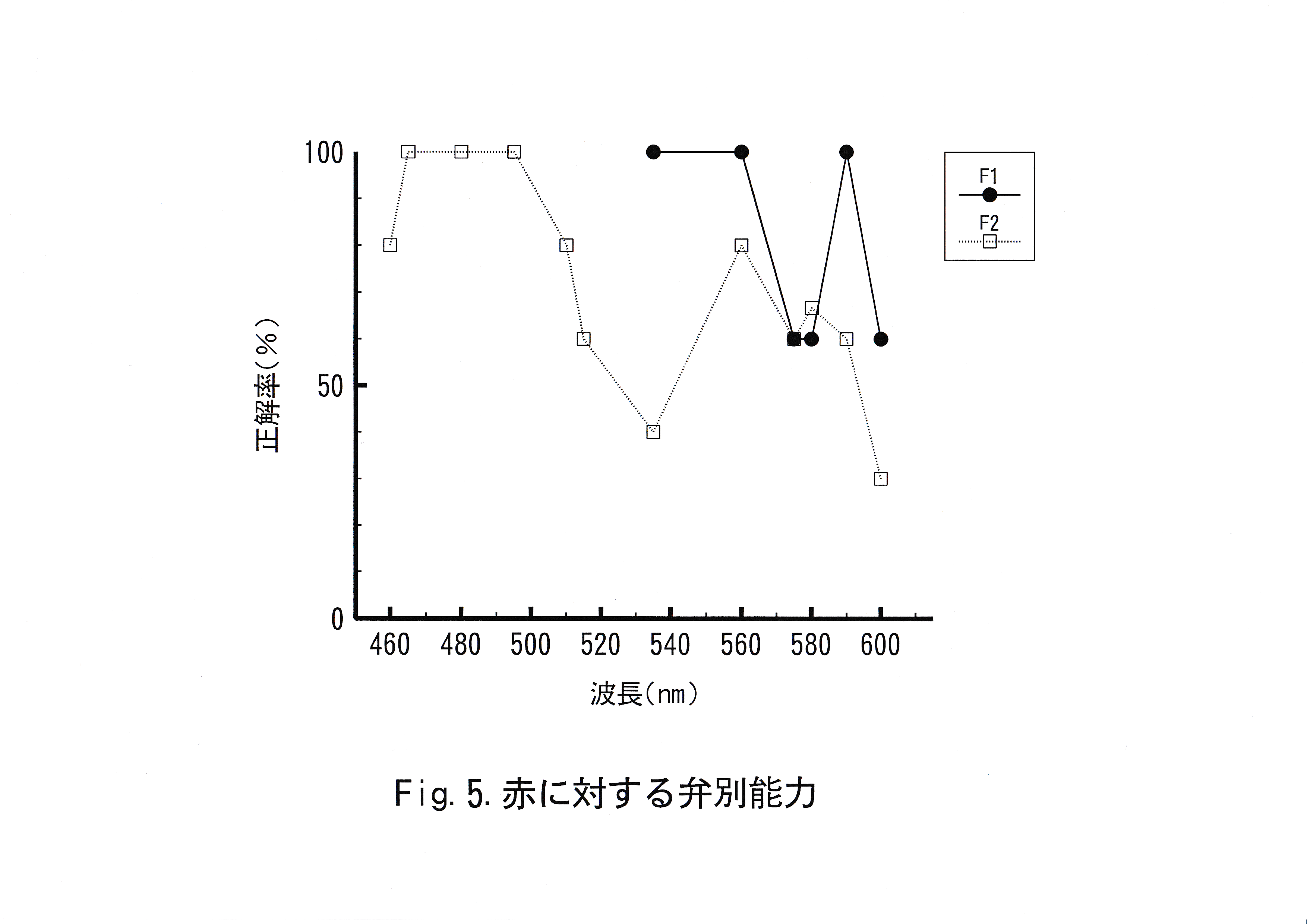
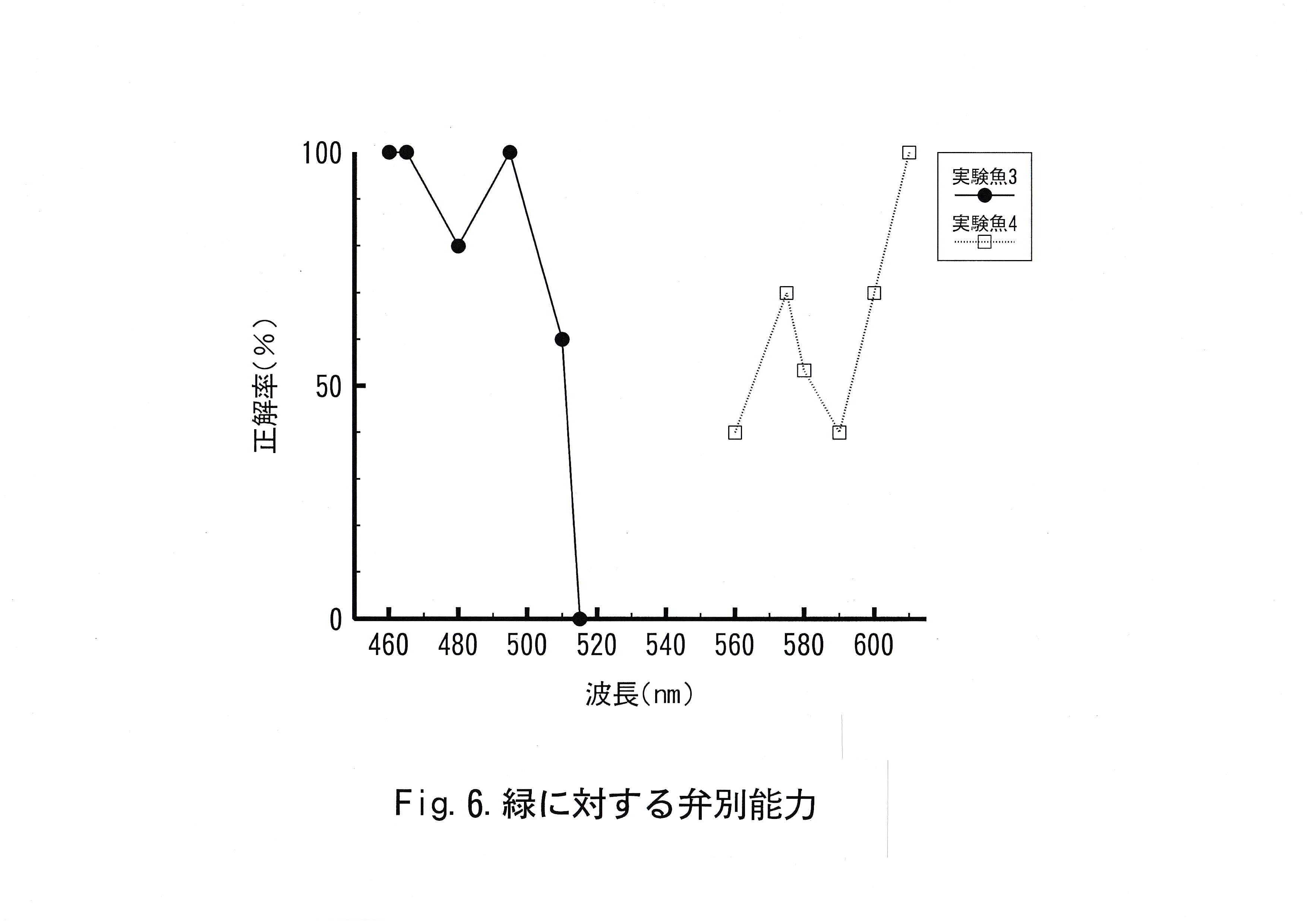
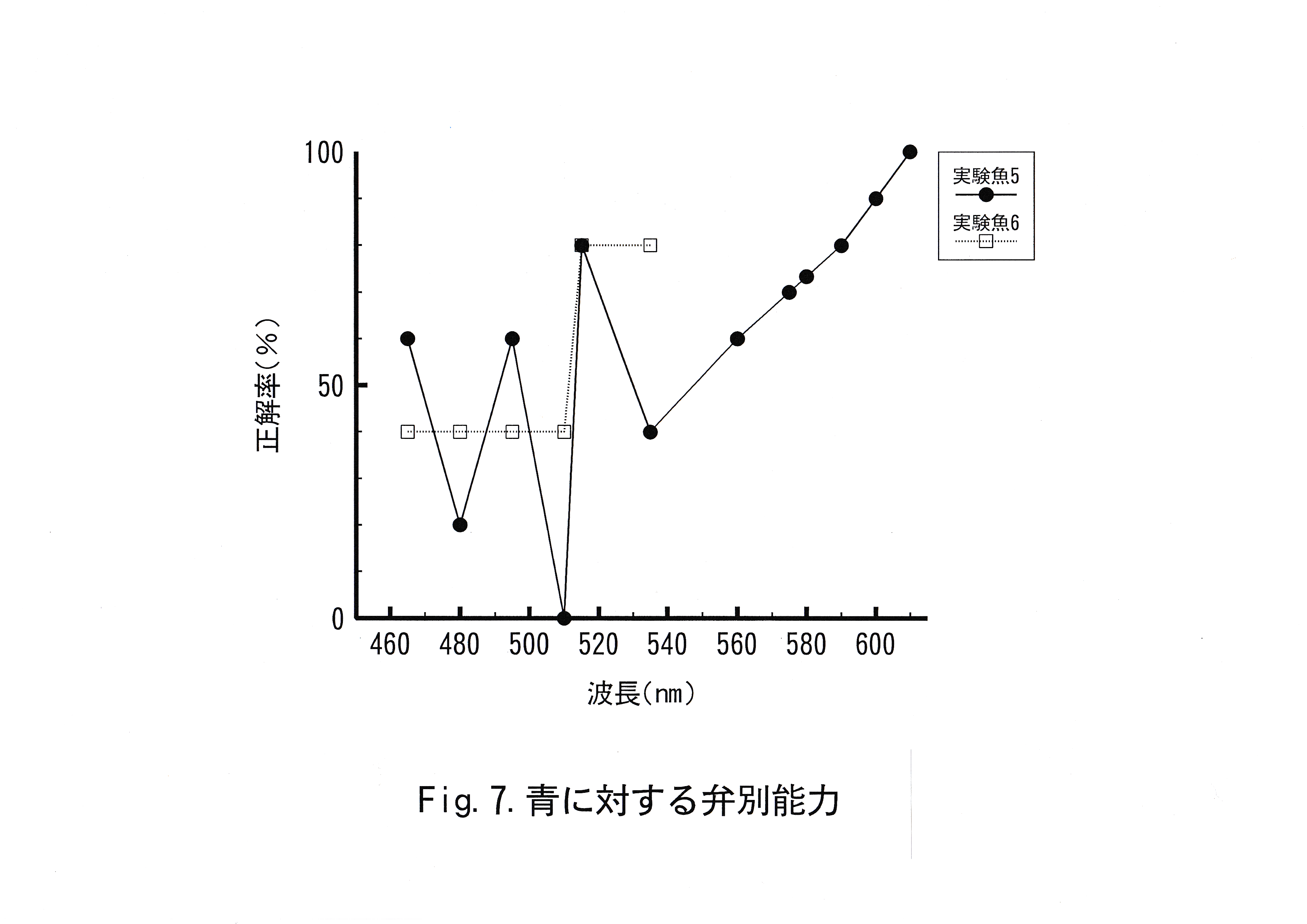
キンギョの赤に対する弁別実験結果をFig.5に示した。キンギョの緑に対する弁別実験結果をFig.6に示した。キンギョの青に対する弁別実験結果をFig.7に示した。これらの図は横軸に波長を示し、縦軸に正解率を示した。実験魚No. 1では波長560 – 590 nm、590 – 600 nmで正解率の低下が認められた。実験魚No. 2では波長495 – 560 nm、560 – 580 nm、580 – 600 nmで正解率の低下が認められた。実験魚No. 3では波長465 – 495 nm、495 – 515 nmで正解率の低下が認められた。 実験魚No. 4では波長560 – 575nm、575 – 610 nmで正解率の低下が認められた。実験魚No. 5では波長465 – 495 nm、495 – 515 nm、515 – 610 nmで正解率の低下が認められた。実験魚No. 6では波長 465 – 515 nmで正解率の低下が認められた。
第3節 考察
実験魚No. 1の結果において、波長590 nmでは正解率100 %をとることから、波長535 – 590 nmの波長域の色は赤と弁別可能と考えられた。赤に対する弁別閾値は波長590 – 600 nmの波長域に存在し、波長差にして20 nm以上あれば赤( 610 nm )と弁別可能と判断された。波長560 – 590 nmの波長域での正解率の低下は、実験魚がこれらの色を正刺激として同一化,すなわち汎化してしまい、赤とは異なる色刺激として差別化、すなわち分化するにはいたらなかった結果と考えられる。本実験では、赤・緑・青に対するキンギョの弁別能力を調べるために、正解率をとった。色彩弁別能力を視力と同様に考えると、正解色と比較する色の波長差が短い場合のみ、正解率が低下することが予想される。しかし、色彩弁別実験では、この波長差が長い場合でも、正解率の低下が予想される。なぜなら正解率の低下には2つの原因がある。1つは、正解率の低下が見られた波長域に弁別閾値が存在し、魚が本当に弁別不可能なことによる正解率の低下である。もう1つは、魚が学習実験では見たことがない色に対して、正刺激に対する行動と同様の行動をとる、いわゆる汎化による正解率の低下である。汎化している色と正解とは区別がつかないというわけではなく、汎化している色を不正解とする学習実験を行った場合は正解率が向上すると考えられる。汎化がなくなることを分化という。
例えば、イヌなどで一定振動数の音に対して条件反射が成立したとき、普通条件刺激を中心とするある一定の振動数域の音が、条件反射を起こさせる(汎化)。この範囲の音域内ではイヌには音の弁別ができないと考えると大変な誤りである。この音域内のある振動数の音と、正しい条件反射音を交互にきかせ、後者の場合には条件反射がおこった直後に強化を行い、前者の場合にはそれをしないことをくり返したとき、その結果として前者にだけ消去がおこれば、その音と条件反射音とはイヌに弁別されていることになる(分化)。このようなテストを汎化音域内の振動数に対して順次行うことにより、真に弁別し得ない範囲を決めることができる(桑原, 1983 )。
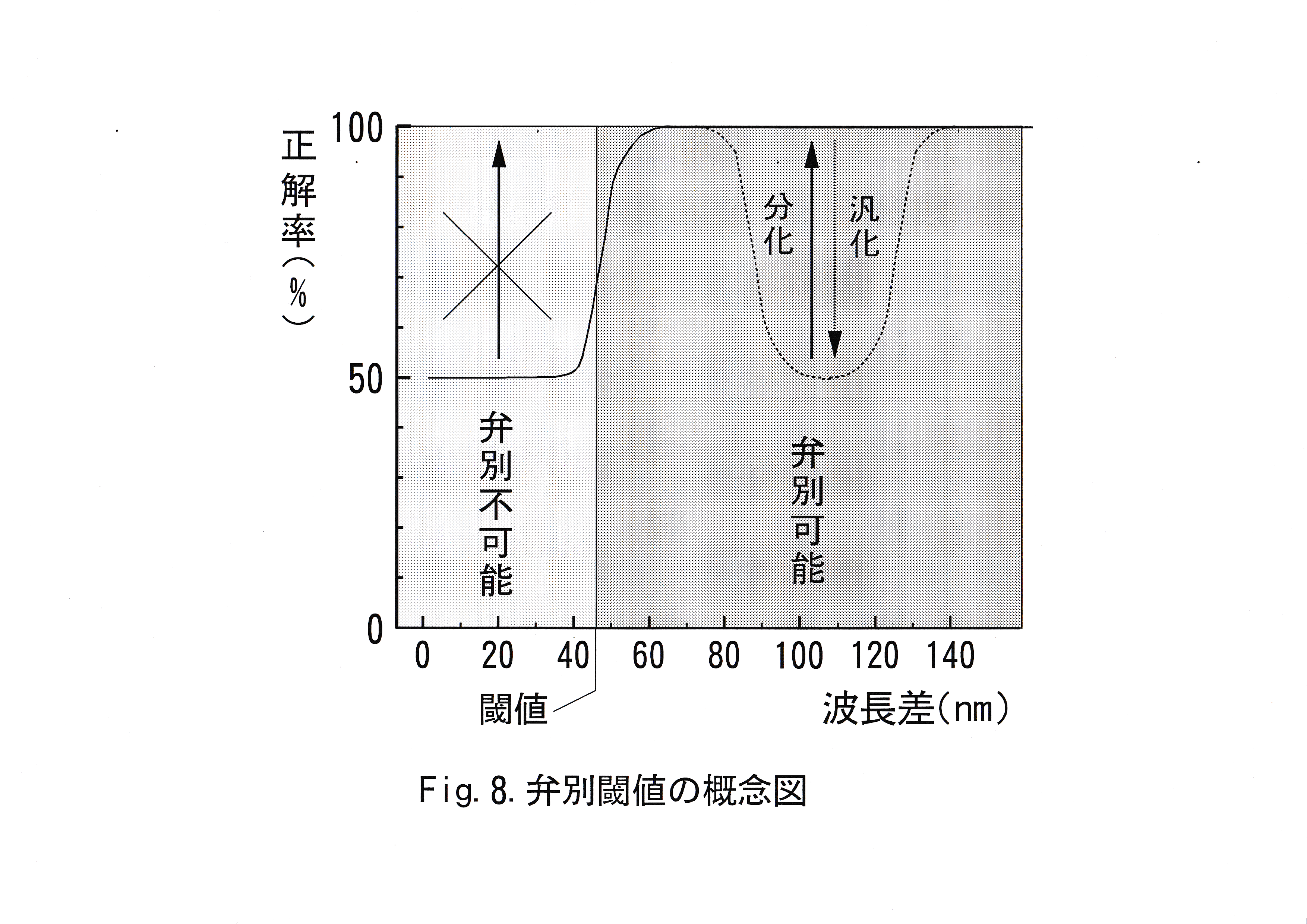
これらのことをふまえ、本実験では、正解率が70 %以上をとる波長の中で、正解色との波長差が最短の波長から、正解色の波長までを弁別閾値が存在する波長域と定義した。Fig.8に弁別閾値の概念図を示す。
実験魚No. 2の結果において、波長560 nmでは正解率 80%をとることから、波長460 – 560 nmの波長域の色は赤と弁別可能と考えられ、赤に対する弁別閾値は波長560 – 600 nmの波長域に存在し、波長差にして50 nm以上あれば赤( 610 nm )と弁別可能と判断された。波長 495 – 560 nmの波長域での正解率の低下は、実験魚がこれらの色を正刺激として汎化した結果と考えられる。
実験魚No. 1と実験魚No. 2にとって本来、同様の実験となる波長 535 – 600 nmの波長域の正解率を比べると、No. 2の方が相対的に正解率が低い傾向にある。正解色から不正解色までの波長差は実験魚 No. 2の方が75 nmも長く、実験魚No. 1は正解色と分化すべき波長が実験魚No. 2よりも短い。波長535 nmにおいて実験魚No. 1の正解率が100 %であるのに対し、実験魚No. 2の正解率が40 %と低いことからも、学習した2色の波長差が短い実験魚No. 1の方が赤と近い波長域の色を赤と分化することができた結果と考えられる。よって、実験魚No. 1の実験結果がキンギョの赤に対する弁別能力としてより特徴的であると判断した。以上のことからキンギョは赤に対する弁別閾値は波長590 – 600 nmの波長域に存在し、波長差にして20 nm以上あれば赤( 610 nm )と弁別可能と考えられる。
実験魚No. 3の結果において、波長495 nmでは正解率100 %をとることから、波長460 – 515 nmの波長域の色は緑と弁別可能と考えられた。緑と緑より短波長側の波長域の色との弁別閾値は波長 495 – 515 nm存在し、波長差にして40 nm以上あれば、緑(波長535 nm )と緑より短波長側の波長域の色は弁別可能と判断された。
実験魚No. 4の結果において、波長575 nmでは正解率70 %をとることから、波長575 – 610 nmの波長域の色は緑と弁別可能と考えられた。緑と緑より長波長側の波長域の色との弁別閾値は波長560 – 575 nm存在し、波長差にして40 nm以上あれば、緑(波長535 nm )と緑より長波長側の波長域の色は弁別可能と判断された。波長575 – 600 nmの波長域での正解率の低下は、実験魚がこれらの色を正刺激として汎化した結果と考えられる。
実験魚No. 3と実験魚No. 4に行った実験の実験条件の違いは不正解色として用いた色が正解色の単波長側か、長波長側かということである。弁別閾値の波長差はともに40 nmであるが、正解率を比較すると、実験魚No. 4の方が実験魚No. 3より、相対的に低い傾向にある。
実験魚No. 5の結果において、波長515 nmでは正解率 80%をとることから、波長515 – 610 nmの波長域の色は青と弁別可能と考えられた。青に対する弁別閾値は波長465 – 515 nmの波長域に存在し、波長差にして55 nm以上あれば青( 460 nm )と弁別可能と判断された。
波長515 – 575 nmの波長域での正解率の低下は、実験魚がこれらの色を正刺激として汎化した結果と考えられる。
実験魚No. 6の結果において、波長515 nmでは正解率80 %をとることから、波長460 – 560 nmの波長域の色は青と弁別可能と考えられた。青に対する弁別閾値は波長465 – 515 nmの波長域に存在し、波長差にして55 nm以上あれば青( 460 nm )と弁別可能と判断された。
実験魚No. 5と実験魚No. 6にとって本来、同様の実験となる波長 465 – 535 nmの波長域の正解率を比べると、No. 5の正解率は、変動が激しくなっている。正解色から不正解色までの波長差は実験魚 No. 5の方が75 nmも長く、実験魚No. 6は正解色と分化すべき波長が実験魚No. 5よりも短い。波長535 nmにおいて実験魚No. 6の正解率が80 %であるのに対し、実験魚No. 5の正解率が40 %と低いことからも、学習した2色の波長差が短い実験魚No. 6の方が青と近い波長域の色を青と分化することができた結果と考えられる。よって、実験魚No. 6の実験結果がキンギョの青に対する弁別能力としてより特徴的であると判断した。以上のことからキンギョは青に対する弁別閾値は波長465 – 515 nmの波長域に存在し、波長差にして55 nm以上あれば青( 460 nm )と弁別可能と考えられる。
今回、正解色とかけ離れた波長域での正解率の低下を汎化によるものと考察したが、これを確認する意味での追加実験を行う必要性が認められた。
第3章 マダイを用いた色相学習実験
第1節 材料と方法
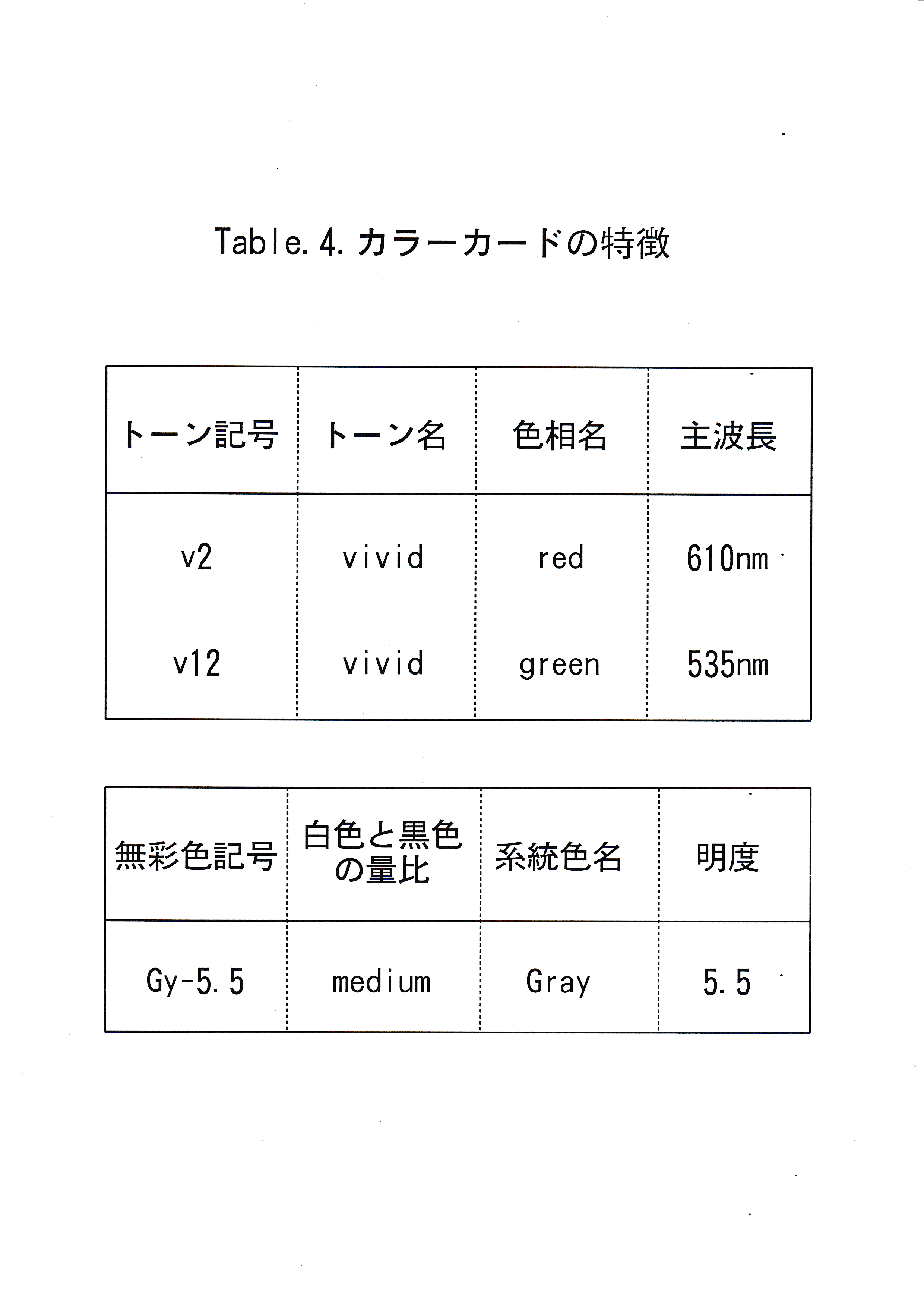
実験魚としてマダイPagrus majorを5尾用いた。実験は東京水産大学、魚群行動学研究室の実験室において、平成7年10月30日から平成7年12月11日まで行った。色刺激として灰色・赤・緑の3枚のカラーカード(日本色研事業株式会社製,新配色カード175b , v2 ・ v12 ・ Gy-5.5 )を6 cm四方に切り、水中に掲示できるように透明プラスチックでコーティング(明光商会社製, L-100 mini MSパウチと、同社製100ミクロンMSパウチフィルムMP 10-90126使用)したカード3枚を用いた。用いたカラーカードの日本色研事業株式会社による色の種類( No. ,色名)をTable.4に示す。 コーティング後のカードの放射輝度を分光放射計(東京光学機械株式会社製,分光放射計 SR-1 )を用いて測定した。
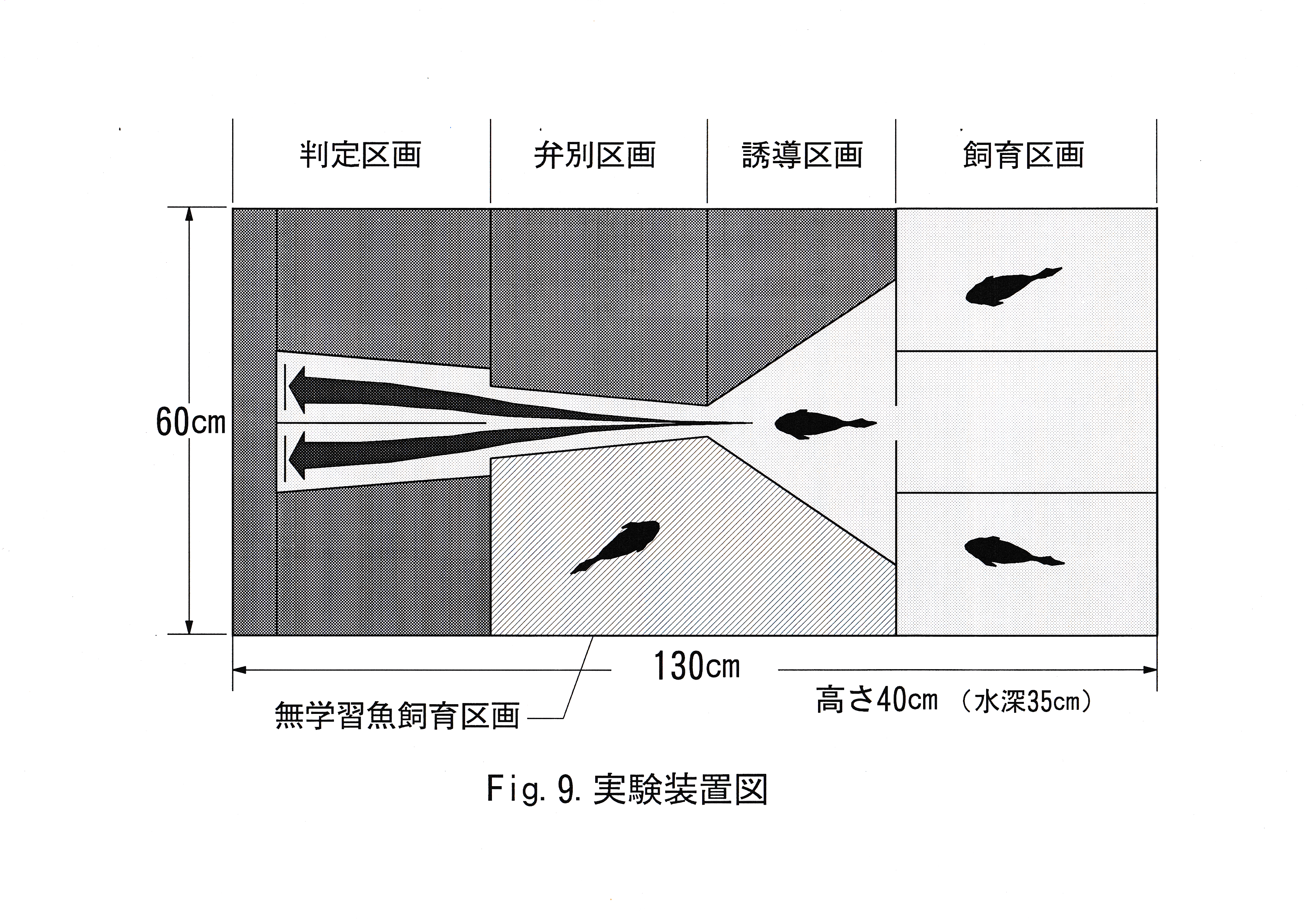
マダイ5尾のうち、1尾はコントロール状態を調べるために用い、3尾は色彩弁別能力を知るために用いた。ここで学習を行わない個体をコントロール魚とし、他の3尾を学習魚と呼ぶことにする。Fig.9に実験装置図を示す。 実験には透明アクリル水槽( 130×60×40 cm )を用いた。海水には人工海水(日本家庭用塩株式会社製,マリンエッセンスN-K-1 )を水道水で溶かしたものを用い、塩分濃度は3.3 – 3.5 ( % )とした。水深は35 cm一定とし、72 l容量のろ過層をもつ濾過装置を用いて濾過し、エアレーションすることにより水質・溶存酸素量を安定させた。サーモスタッド・ヒーターを使用し、飼育時の水温を25℃一定とした。これらの装置は飼育時・実験時ともに水槽内に設置したが、極力実験魚に視認されることのないよう配慮した。水槽内に黒の塩ビシートを貼り付け、実験魚に色刺激以外の色覚的要素を認識させないようにした。水槽内をアクリル板で仕切ることにより、飼育区画・誘導区画・弁別区画・判定区画の4つの区画を作製した。それぞれの区画の間にはゲート(開口部の高さ19cm)を作製し、遠隔操作により開閉できるようにした。飼育区画は仕切により、さらに3つにわけられ、学習魚を1尾ずつ収容できるようにした。誘導区画は実験魚に無駄な行動をさせないための区画で実験はこの区画に実験魚を収容した状態から始めた。判定区画は実験魚が色を弁別したかを判定するための区画で、中央の仕切により、左右にわけられ、実験魚が弁別区画からゲートを通過する際に、区画の左右どちらかにしか侵入できないようにした。区画の左右に1枚ずつカードを掲示する掲示場所を作製した。掲示場は誘導区画から弁別区画に侵入しようとする魚が左右2枚のカラーカードを同時に視認できる位置に設置した。カードを色刺激として学習しやすいようにカード掲示場所の真下に給餌口を作成した。弁別区画は細長い水路状になっており、実験魚がここを通過する際、判定区画の左右どちらへ入るかを選択する場となっている。
餌として沈降性のドライペレット(坂本餌料株式会社製,マダイ稚魚クランブルC4 )を1尾について1日60粒与えた。餌は実験開始前にカードを掲示するアクリル板上方の給餌装置の中に3粒ずつ入れておく。給餌の際は暗幕の外から遠隔操作によって給餌装置を動かし餌を落とす。餌の落ちる位置に漏斗があり、餌は漏斗と塩ビパイプでつながっている給餌口から出てくるようになっている。
実験水槽は暗幕内に置き、20wの蛍光灯(岩崎電気株式会社製, アイライン,白色, FL20SW ) 2本を照明器具内に収容したものを照明として用いた。水槽全体を照らすために照明器具を水面中央から高さ60 cmの位置に設置した。カード掲示位置の水中照度は159 lxであった。照明点灯後魚の網膜を明順応状態とするために1時間以上順応させた後に実験を開始した。
実験時には飼育区画と誘導区画の間のゲートを開放し、誘導区画内に魚が侵入したのを確認した後にゲートを閉じ、観察者は実験結果に影響しないように暗幕の外に退去した。暗幕内にCCDカメラを設置し、実験観察は暗幕外のモニターで行った。
学習魚は実験時以外は飼育区画内に収容し、実験時にのみ、判定区画内の給餌口にて給餌させた。コントロール魚は実験時以外は区画外で飼育・給餌した。コントロール魚に対しては、判定区画内にカードを掲示しない状態で実験を行った。実験魚を刺激しないように誘導区画内へ移動させた後に、判定区画と弁別区画の間と、弁別区画と誘導区画の間のゲートを開放し、自由遊泳させた。実験魚が判定区画に進入した際に、左右どちらに進入したかを1000回記録した。コントロール魚に対してはカードの掲示を行わなかった。学習魚に行う実験ではカードの掲示方向は最初は正解を右とし、正解したら左右逆とすることから、この方法で正解・不正解の方向を決めた。進入20回を1実験とし、実験回数ごとに正解率を求めた。正解率は進入回数に対する正解の割合と定義した。
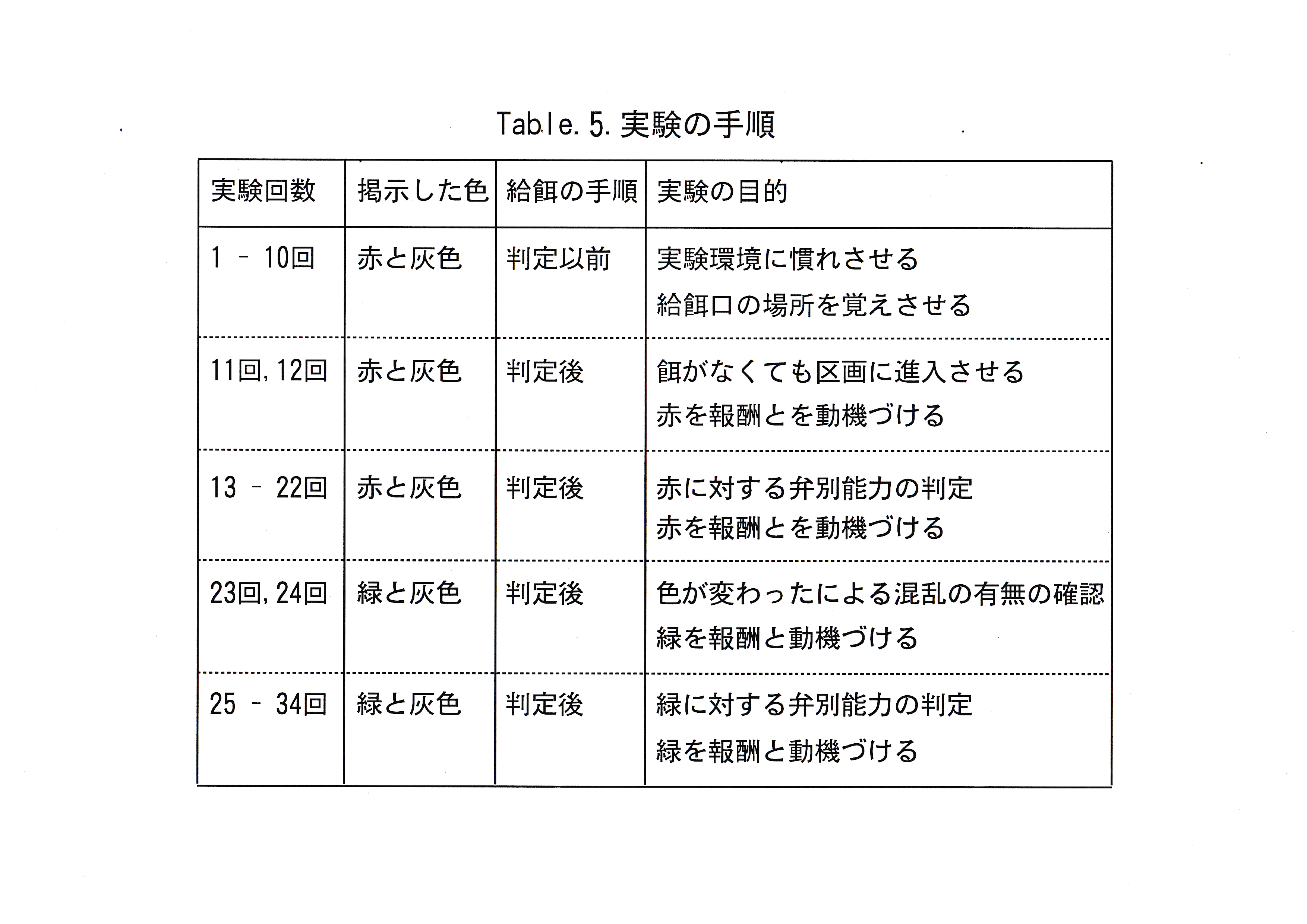
学習魚にはカード掲示場に赤( v2 ) ・緑( v12 )のうち一方を正解とし、灰色( Gy-5.5 )を不正解として掲示し、正解の場合は報酬として餌を与え、不正解の場合は報酬を与えられないという学習を1日20試行施した。20試行の学習を1実験とし、3尾ずつ計34回の実験を行った。実験の手順をTable.5に示した。 まず赤と灰色のカードを掲示して実験を行った。実験10回目までは餌が給餌口にあるという条件で実験を行い、実験11回目以降は実験魚が判定区画内に進入してから給餌した。実験魚が実験手順を覚え、2つの区画を見比べる行動が3尾で観察されたのが実験12回目であったので、実験13回目から22回目までの実験10回を赤に対する弁別能力の判定実験とした。続けて正解色を緑にして同様の実験を行った。2つの区画を見比べる行動が実験魚3尾で観察されたのが実験24回目であったので、実験25回目から34回目までの実験10回を緑に対する弁別能力の判定実験とした。実験魚の判定には統計学的手法を用いた。コントロール魚の実験20回の正解率の分布の歪度・尖度による正規性検定を行った。正規性がない場合は対数変換により、データを正規分布に近づけた。正解率の分布に正規性が認められた場合は以下の推定を行った。まず、信頼係数99.9 ( % )でコントロール魚がとりうる正解率の母平均μ( % )を区間推定した。推定した最大のμ以上の正解率を全実験回数中、最大何回とりうるかという母比率p ( % )の信頼係数99.9 ( % )の区間推定を行った。これらの推定を行った後にコントロール魚の正解率の平均値と弁別率を算出した。弁別率はコントロール魚がとりうる最大の正解率の母平均μ( % )以上の正解率をとった実験回数の、全実験回数に対する割合と定義した。学習魚3尾の赤と灰色に関する判定実験10回の正解率の平均値をとり、弁別率を算出した。同様に緑と灰色に関する判定実験10回の正解率の平均値をとり、弁別率を算出した。
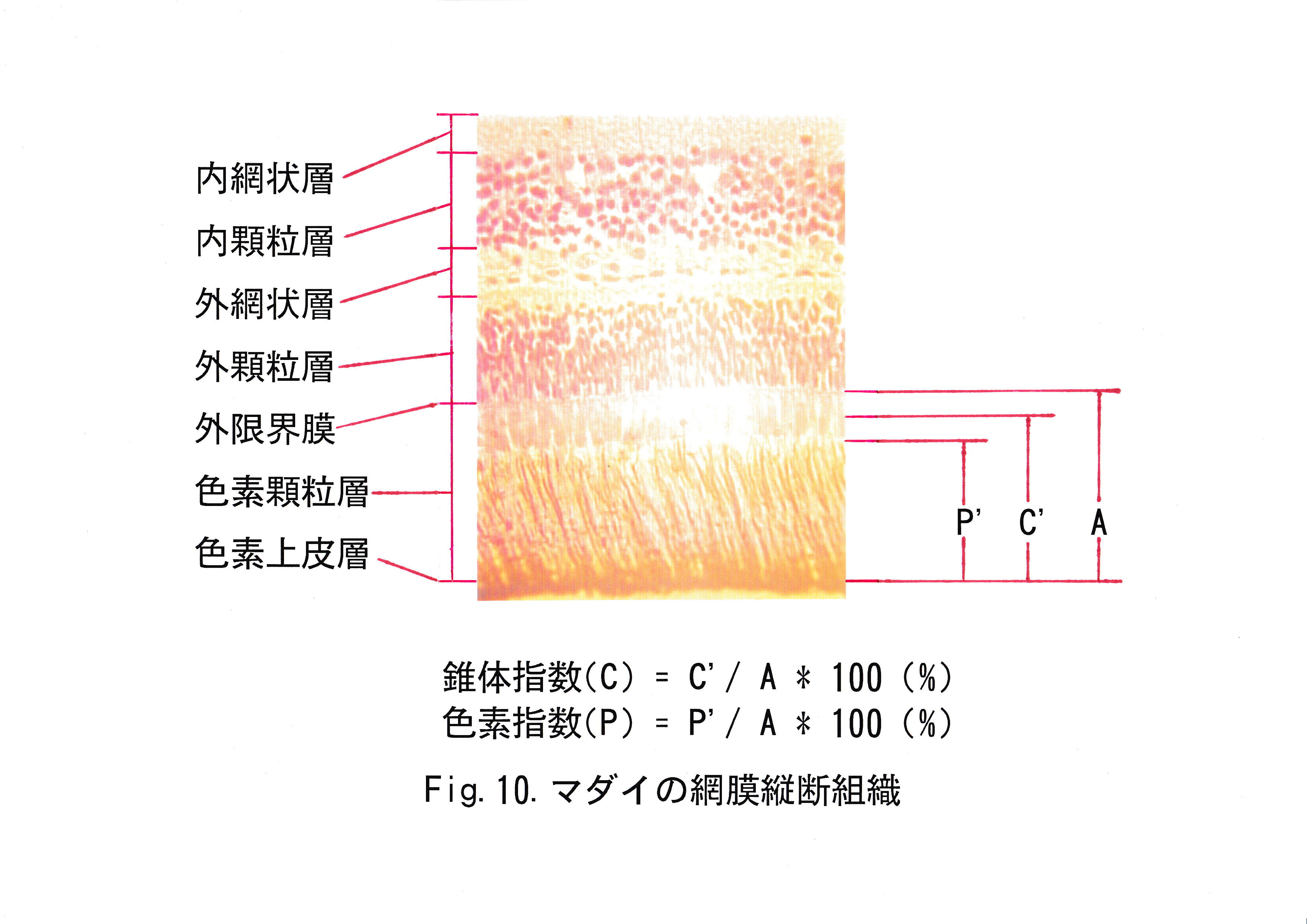
網膜の順応状態を知るためにマダイ1尾を用い、網膜縦断組織切片を作製した。マダイを水槽内に1日収容し、照明点灯1時間後の網膜をブアン液を用いて固定した。固定した網膜はアルコール脱水、キシレン透徹、パラフィン包埋、ミクロトームによる組織の縦断、 ヘマトキシリン・エオシン(HE)染色、封入剤(MGK-S)標本封入という作業過程を経てプレパラートにした。これを顕微鏡で顕鏡した後、ビデオプリンタに出力した。縦断組織切片の顕鏡例をFig.10に示す。
西島( 1995 )に従い、網膜切片について光に対する明順応状態の順応率を表す指標として、錐体指数( C ) ・色素指数( P )を用いた。色素上皮層から外限界膜までの幅をAとし、色素上皮層から錐体の楕円体の中心までの幅をC’、および色素顆粒層上縁部までの幅をP’として、それぞれの長さを測定した。
錐体指数( C )はAに対するC’の割合とし、色素指数( P )はAに対するP’として、それぞれ百分率で求めた。
錐体指数( C ) = C’ / A * 100 ( % )
色素指数( P ) = P’ / A * 100 ( % )
第2節 結果
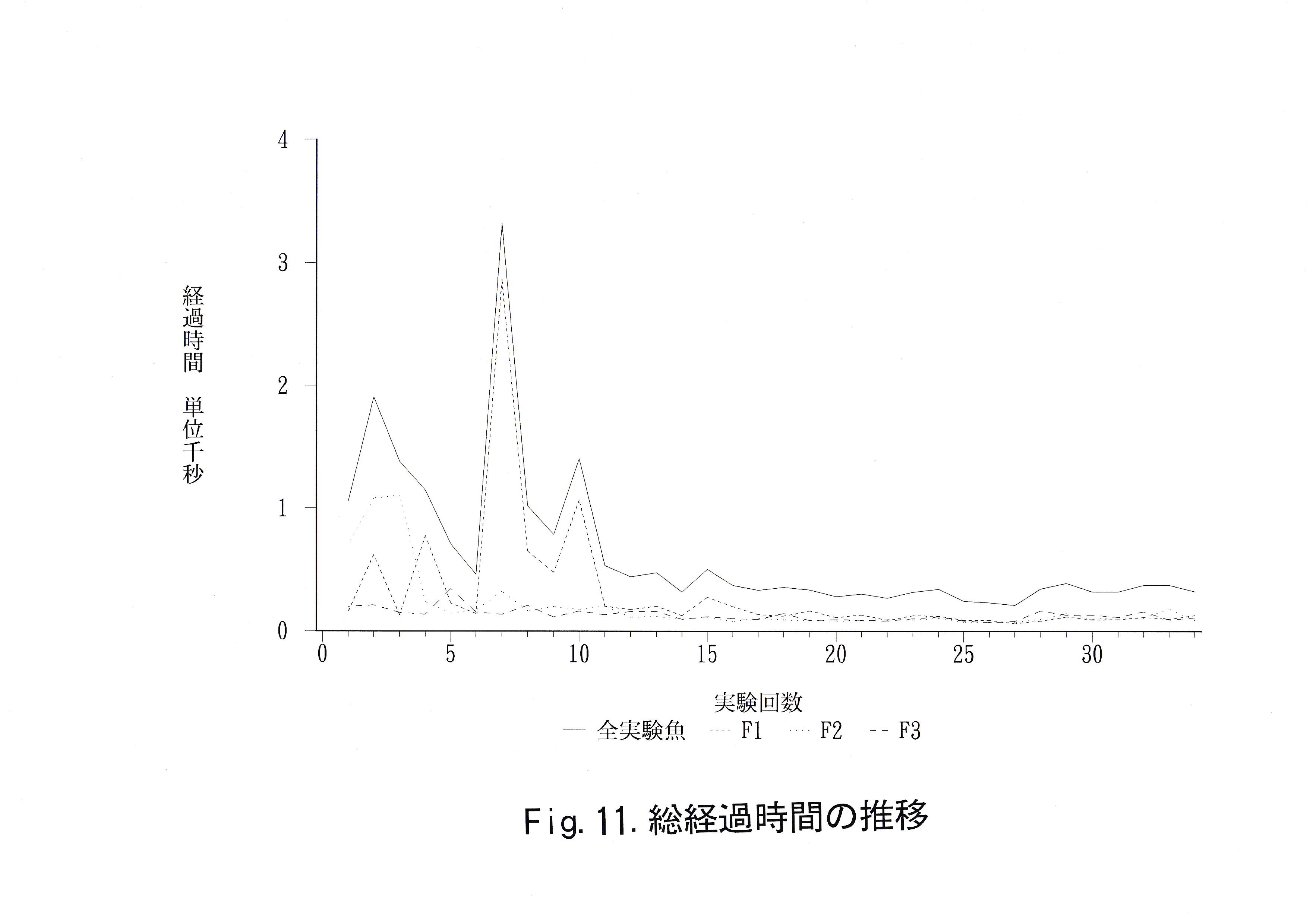
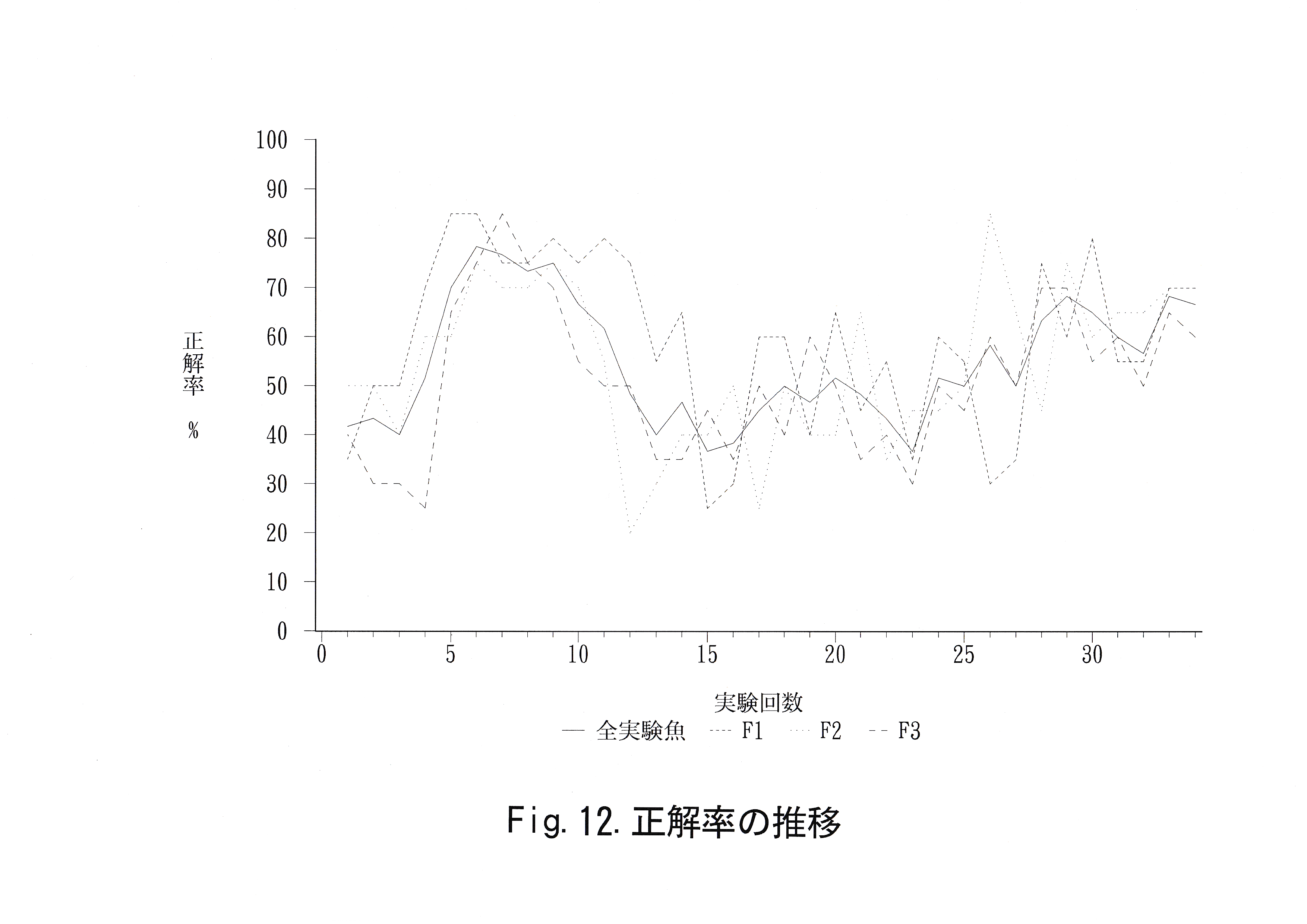
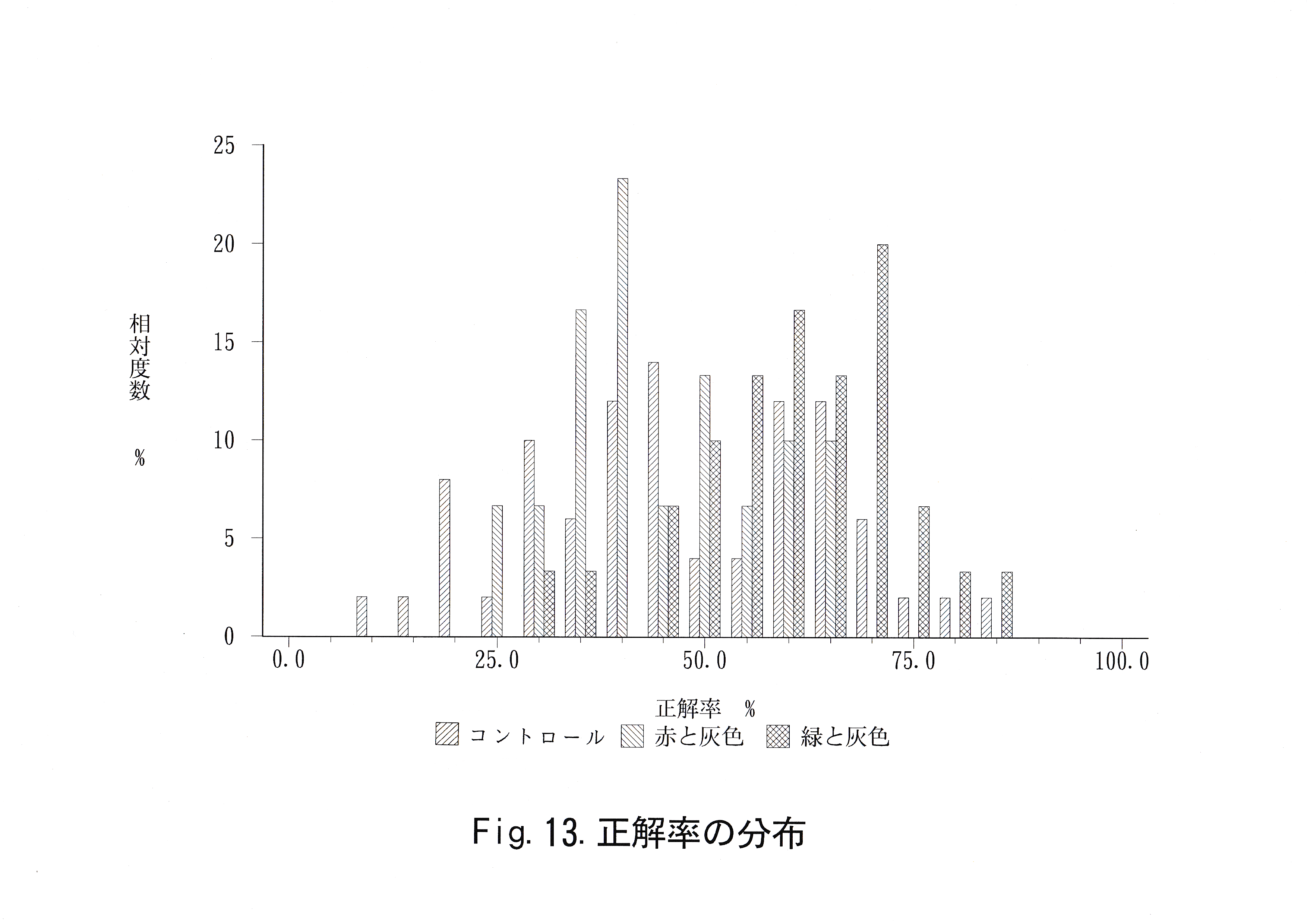
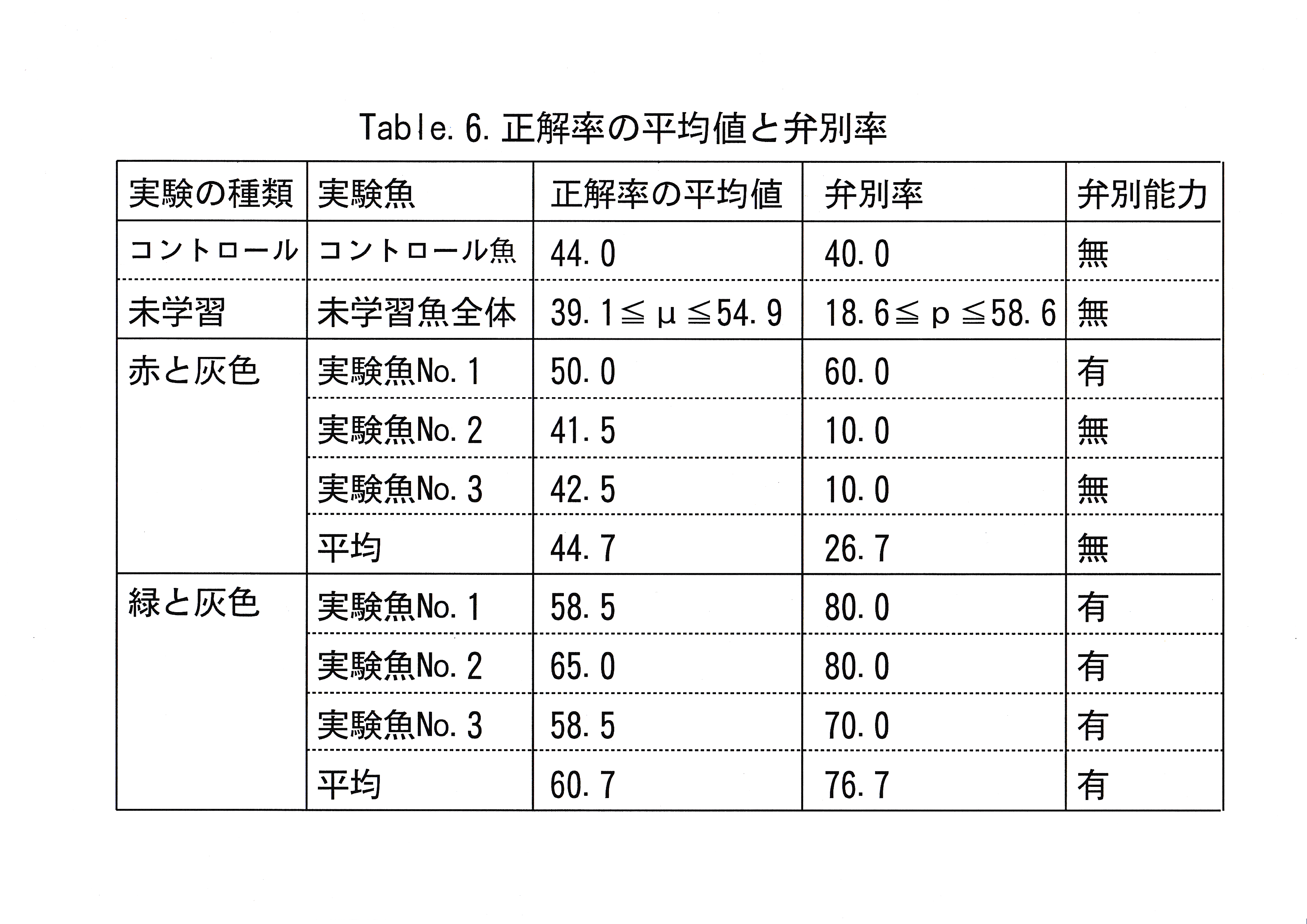
実験回数に対する総経過時間の推移をFig.11に示した。 実験回数に対する正解率の推移をFig.12に示した。 無学習魚を用いて得られた正解率、学習魚を用いて得られた赤・緑に対する正解率の分布をFig.13に示した。 コントロール魚のデータから推定した未学習魚の正解率の母平均、その母平均の上限を超える正解率をとる実験回数の母比率とコントロール魚・学習魚の正解率の平均値、弁別率をTable.6に示す。
コントロール魚の正解率の分布には正規性が認められた。信頼係数 99.9 ( % )で未学習魚がとりうる正解率の母平均μ( % )は 39.1≦μ≦54.9( % )であった。未学習魚が平均としてとりうる限界の 正解率を55( % )として、55 ( % )以上の正解率を全実験回数中、最大何回とりうるかという信頼係数99.9 ( % )の母比率( p )は 18.6≦p≦58.6 ( % )であった。コントロール魚の正解率の平均値は 44 ( % )であり、弁別率は40 ( % )であった。
赤と灰色に関する判定実験10回の正解率の平均値は実験魚1が 50.0 ( % )、実験魚2が41.5 ( % )、実験魚3が42.5 ( % )、全体で44.7 ( % )であった。弁別率は実験魚No. 1が60.0 ( % )、実験魚No. 2が10.0 ( % )、 実験魚No. 3が10.0 ( % )、全体で26.7 ( % )であった。緑と灰色に関する判定実験10回の正解率の平均値は実験魚No. 1が58.5 ( % )、 実験魚No. 2が65.0 ( % )、実験魚No. 3が 58.5 ( % )、全体で60.7 ( % )であった。弁別率は実験魚No. 1が80.0 ( % )、実験魚No. 2が80.0 ( % )、実験魚No. 3が70.0 ( % )、全体で76.7 ( % )であった。
錐体指数( C )と色素指数( P )は以下の通りであった。
錐体指数( C ) = C’ / A * 100 ( % )
= 2.98 / 3.35 * 100 = 88.96 ( % )
色素指数( P ) = P’ / A * 100 ( % )
= 2.60 / 3.35 * 100 = 77.61 ( % )
第3節 考察
赤を正解とする実験の正解率の平均値はいずれも未学習魚がとりうる正解率の母平均μの範囲内であり、弁別率で見ても、2尾は正解率の母比率pの範囲を下回り、pを上回った実験魚No. 1においてもpの上限を1.4 ( % )上回る程度のことから、マダイは赤と灰色を弁別することが不可能と考えられ、仮に弁別可能であったとしても困難であることが示唆された。さらに今回の実験から、マダイが赤を知覚しないことを確認する意味での追加実験を行う必要性が認められた。
緑を正解とする実験の正解率の平均値はいずれの実験魚も未学習魚がとりうる正解率の母平均μの上限を上回り、弁別率で見ても、全実験魚が正解率の母比率pの上限を18.1 ( % )も上回ることから、マダイは緑と灰色を弁別可能であると考えられた。
錐体指数( C )・色素指数( P )が大きくなるほど明順応状態に近づき、小さくなるほど暗順応に近づく。これらがともに77 ( % )以上と算出されたことから、本実験での照明下で実験魚の網膜が明順応状態となっていることが確認された。
第4章 総合考察
第1節 総合考察
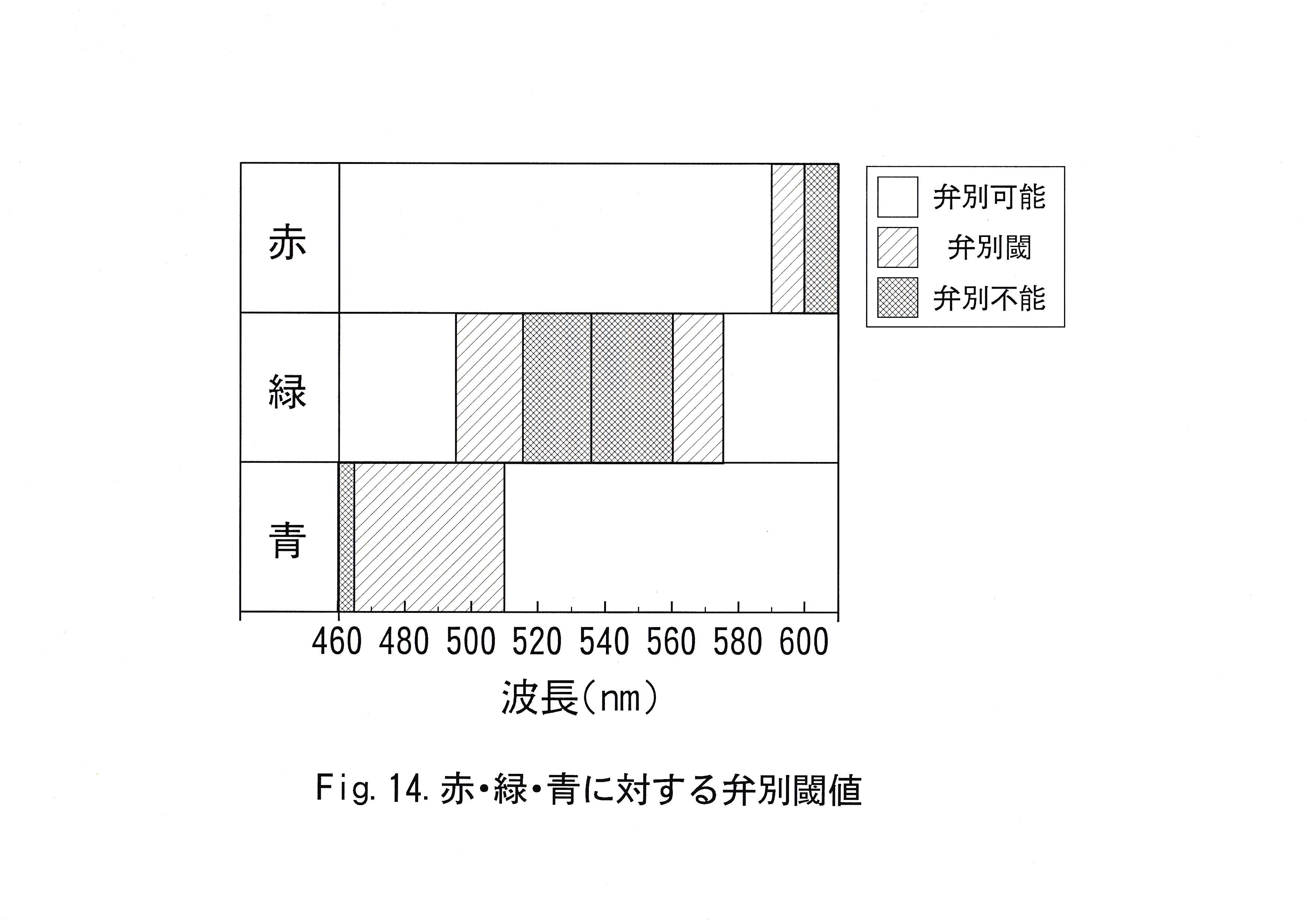
本研究において「キンギョは物体色の赤・緑・青を弁別できるのか」、「弁別可能ならばその閾値はどのくらいなのか」「学習あるいは弁別させる色の組み合わせにより結果に違いがないか」ということついて調べた。キンギョは赤・緑・青を弁別可能であることが確認できた。赤・緑・青に対する弁別閾値を模式的にFig.14に示す。 学習・弁別させる色の組み合わせにより弁別閾値には差が見られた。弁別閾値は赤(波長610 nm )に対して波長差20 nm、緑(波長 535 nm )に対して40 nm、青(波長460 nm )に対して55 nmに存在し、弁別能力は長波長に対して高く、短波長に対して低いと考えられた。
キンギョの網膜中のλmax= 455±15 nm (青)、λmax= 530±5 nm (緑)、λmax= 625±5 nm (赤)の異なった色素を有する3種の錐体が存在するという報告( Marks , 1965 )がある。
キンギョの青に対する感光色素の波長特性が他の色素と比較して明らかなピークをもつものではないことと、本実験でキンギョの弁別閾値が短波長の色に対して低いとされたことの関係の有無を知るための実験が必要と考えられた。
マダイの採餌活動は吸収の最大値が430 – 500 nmの色彩物質によって誘導されること(伊奈ら, 1979 )や、マダイが青色( 450 nm付近)と黄色及び赤色( 600 – 700 nm付近)の色相を好むこと(柿元ら, 1995 )が報告され、マダイのスペクトル感度曲線が470 nmで最大値をとり、550 nmと600 nmで副次的な最大値をとることから、青に敏感な要素が著しく高く、約3 lxの弱光に順応した眼から525 nmにピークをもつ感度曲線が得られ、 650 nmより長波長の光に網膜がほとんど感じない( Kobayashi , 1962 )とされている。さらにマダイのL型S-電位は522 nm付近にピークがあり、波長633 nmの光に対する網膜感度はかなり低いと報告されている(川村ら, 1991 )。
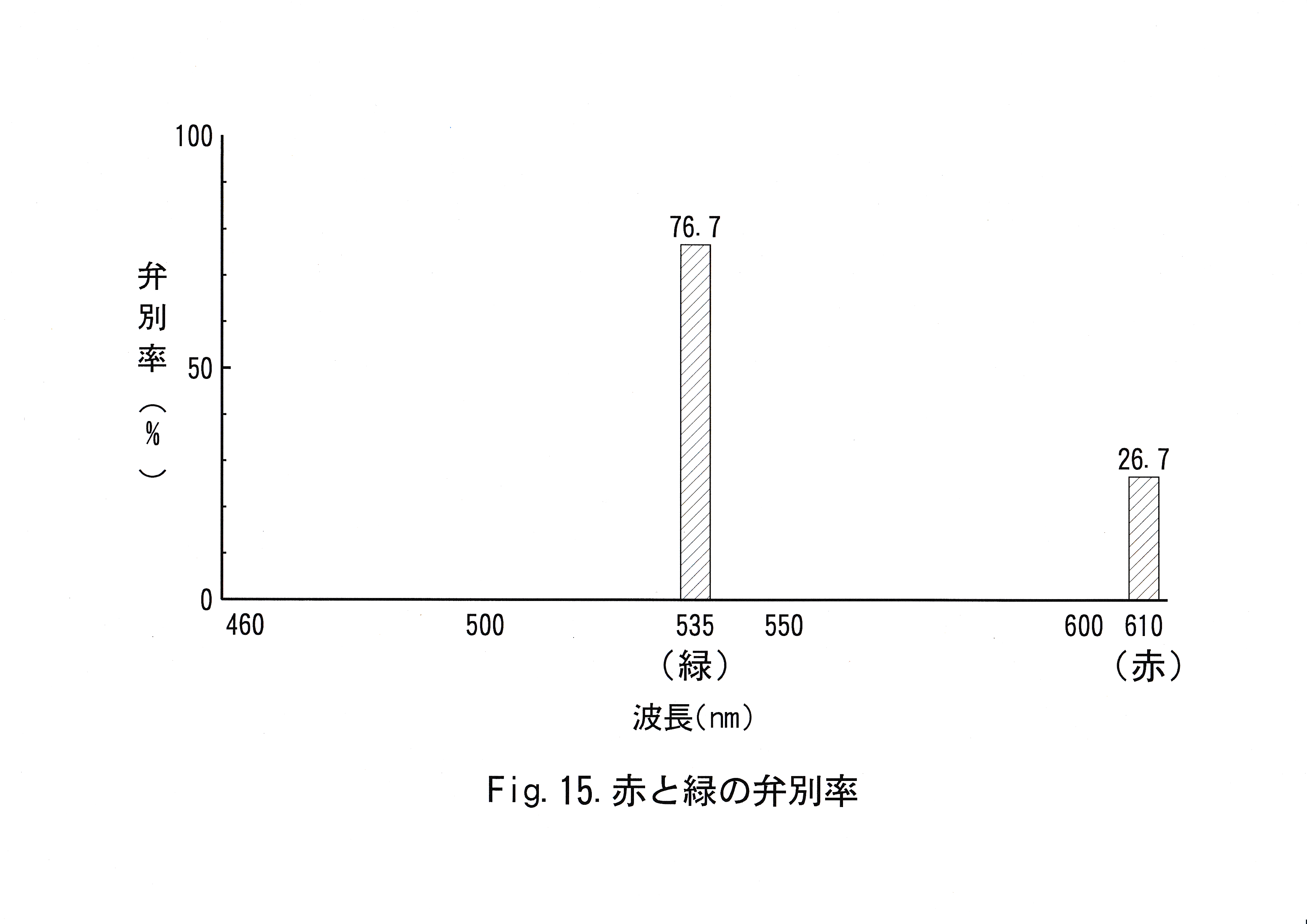
本研究において、マダイは赤(波長610 nm )と灰色を弁別することが不可能と考えられ、仮に弁別可能であったとしても困難であることが示唆された。さらにマダイは緑(波長535 nm )と灰色を弁別可能であると考えられた。Fig.15に赤と緑の弁別率を示す。 マダイは長波長側の色に対する弁別能力が低いと考えられるが、これを確認するためにも、行動学的にマダイの波長ごとの視感度を得られる手法が必要と考えられる。
第2節 実験の問題点
キンギョの学習・弁別実験を一通り終えたところで、今回の実験設定に問題があると考えられた。まず、色とは波長差に従って段階的に変化するものではないということである。今回のキンギョを用いた実験では、正解色と比較色を学習させ、比較色の波長を正解色に近づける際にキンギョにとって正解色とも比較色ともいえない色が存在している場合があった。ヒトの色覚では反対色説に基づいて、ユニーク色と分類される青・緑・黄・赤がある。これらの色は人の目に異なる色彩を感じさせる。例えば信号は青(緑) ・黄・赤が1色だけ点灯するが、われわれが黄色を見たときに、黄色が青と赤のどちらに近い色と感じるだろうか。おそらくどちらの色にも近いとは認識しないであろう。
今回の実験では3色説に基づいて、赤・緑・青で色を代表した。なぜ赤・緑・青なのかというと、この3色を用いることにより心理物理的には全ての色が表現できるからである。しかしヒトの場合は赤から青の波長域には大きく分けて赤・黄・緑・青の心理的4原色(ユニーク色)を感じ、弁別実験で用いたカラーカードをヒトが見ると17の色相に区別できる。学習実験では実験魚1尾に対し赤・緑・青うちの2色ずつ6通りで学習させた。キンギョがヒトと同様の色相を有する場合、例えば学習実験で赤を正解、緑を不正解とし、弁別実験で赤と黄色を見せた魚には赤・緑のどちらとも認識しない色を見せてしまい、黄色と赤を区別できながら、黄色に対して赤に対する行動と同様の行動をとらせてしまったわけである。
この実験が無意味であるかというとそうではない。赤・緑・青以外の色相をキンギョが認識するとき、その色がどの波長域に存在するかを知るのに、今回行った用いた実験が有効と考えられる。ただし、魚が弁別閾値とは無関係の波長域で正解率が低下するときに、その波長の色を本当は魚が弁別できることが証明できなければならない。
今回の実験で汎化によると仮定した正解率の低下が見られた波長域の色に対しては、実験魚が弁別可能であることを確認する実験が必要と考えられる。またこのような汎化がおこった原因の一つとして、その色が実験魚に誘引効果をもつか否かを調べる実験も必要と考えられる。
マダイの実験の設定においてマダイの見る色の明るさが赤・緑・灰色で等しいことが大前提になる。しかし今回の実験では異なる明るさの灰色を複数用いて比較するということは行わず、ヒトの比視感度において明るさが等しいとされる灰色を用いているためマダイにとっては色の明るさが異なる可能性がある。赤と灰色が弁別できるのならその明るさを実験魚が認識した可能性がある。しかし弁別できなかったことからはマダイが色の違いとともに明るさの違いを認識していないことを示す。その後、赤との明るさに差がない緑に対して弁別可能であることから、マダイの色彩弁別能力に対する判断は妥当かと思われる。しかし、今回の設定には光源とその照度、色刺激そのものについて改良すべき点が多数あり、今後は色覚を有するか否かを調べる手法を確立する必要があると考えられる。
第3節 魚類の色覚研究
魚類は一般によく発達した構造の眼をもっており、餌をとり、仲間と群を作り、あるいは敵から逃げるという多様な生活のなかで活用されている(有元, 1989 )。視覚器としての眼の働きは、光の質的、量的な差異を認識することであり、これが感覚として認知されている。視覚が光の量的な差異の認識(認知)であるなら、光の質的な差異の認識(認知)は色覚であろう(張, 1993 )。
魚類に限らず、動物は太陽エネルギーを電磁波として利用している。ガンマ線、エックス線、紫外線、赤外線、マイクロ波、ラジオ波などを一般に電磁波というが、ものを見るために必要な光も本質的には電磁波とかわらない。ただ電磁波の波長範囲は10^-10 cm程度のガンマ線から、ラジオ波ともなるとガンマ線の波長の10^16倍程度の数kmの長さにもなるが、ヒトおよび魚の眼に見える範囲、つまり可視域はせいぜい380nmから760 nm ( nanometerの略で1 nm = 10^-9 m = 1 mμ)である。水中生活をしている魚にとっては利用しやすい光線は、水が透過した光線だけなので、水が光線を選択吸収する事実を考える必要があり、また海水の含む塵埃や浮遊生物が光線の透過を妨げることを考える必要がある。魚類における視覚研究は摂餌、配偶、攻撃、逃避、群形成、縄張り形成などの行動学的諸問題、魚類の誘致(集魚灯、擬餌など)、魚の行動の制限(定置網における垣網など)、魚の生息条件の改善(築磯など)の漁労・増殖上の諸問題とも大なり小なり関係する。それ故に、魚類の色覚に関しては、より一層広範囲の説明や考察が必要である(丹羽, 1991 )。
従来色彩感覚の研究の多くは学習実験による手法がとられていたが、電気生理学、顕微分光光学的研究手段の導入により、魚類の色覚に関する研究が一段と進み、魚類の視覚機能の解明に今後も大きな役割を果たすものと認められる(張, 1993 )。近年、魚類の色覚神経系における光環境への適応進化過程を考察し、これを工学システムの最適化モデルの研究に応用しようという試みがある(袋谷, 1991 )。
色覚について錐体では三色説、水平細胞から中枢神経系において反対色説が成り立つという段階説が現在有力である(大山, 1994 )。生理学的手法や顕微分光測定法の技術的進歩により、これらの結果は色覚が段階説に則ることを裏付けるものとなっている(張, 1993 )。色刺激が網膜で受容されてからの情報の伝搬を色感覚と言い、その情報が脳に到達後、最終的に色と知覚させる感覚を色知覚という。色感覚については段階説による説明が可能だが、色知覚については未だわかっていないことが多い。色知覚を知るには電気生理学的手法を用いて、脳内の色の受容野を決定し、脳内神経電位を比較する方法が考えられ、その手法の確立が急務と考えられる。
一方、行動学的手法を用いて、魚の色覚の有無を知ることは、色刺激の設定が十分であれば可能と考えられる。色を示す物理量は現在波長しかないが、色覚を標準観測者の色覚をもとに心理的物理量で示す試みは行われている。魚類の色覚を説明するには、この標準観測者と比較して説明することが考えられ、そのためには行動学的手法をより客観性のあるものにする必要がある。
これらの魚類の視覚機能に関する研究が進歩すれば、さらに魚類の視覚を漁獲技術に応用することが可能と考えられる。
謝辞
本研究をまとめるにあたり、魚類の視覚生理的機能や行動学的研究手法に関する博識だけでなく、研究に対する姿勢を教えて頂いた魚群行動学研究室有元貴文教授に謹んで感謝の意を表します。
実験の重要な材料である色刺激にカラーカードを利用するという発想を頂き、本研究を通して多大な御助力を承った同研究室秋山清二助手に深く感謝の意を表します。実験観察に必要不可欠であった撮影機材を快く貸して頂き、電子機器等に関する知識を絶えず授けて頂いた同研究室田原陽三顧問に深く感謝の意を表します。
実験魚の飼育や実験機材の作成に必要な知識、その他の博識を授けて頂いた同研究室博士課程角田篤弘氏に深く感謝の意を表します。本研究を通して、魚類の視覚機能に関する研究の師匠として終始御相談にのって頂き、研究者としての素養を授けて頂いた同研究室博士課程塩原 泰氏に深く感謝の意を表します。電算機や光学に関する知識や自然科学に法則性を見出す姿勢を教えて頂いた同研究室博士課程崔 淅珍氏に深く感謝の意を表します。電算機に関する知識や基礎研究の重要性と困難、研究者に必要な人間性を教えて頂いた同研究室修士課程韓 斌氏に深く感謝の意を表します。研究を様々な視野から考えることや、研究発表への取り組み方を教えて頂いた同研究室修士課程渡邊尚明氏に深く感謝の意を表します。統計学的手法や実験設定に関する御助言を頂いた同研究室修士課程アリプルバヤント氏に深く感謝の意を表します。研究の困難に対して本人の身になって相談にのっていただいた同研究室修士課程五十風純氏に深く感謝の意を表します。最後に本研究に対し深い御理解と有益な御助言を賜った水産工学研究所張 秀梅博士に深く感謝の意を表します。
引用文献
有元貴文( 1989 )
魚類の生態からみた漁法の検討
24.魚はどうして光に集まるのか (下)
水産の研究, 8巻, 5号( 38 ), p.39
Bowmaker, J. K., A. Thorpe and R. H. Douglas ( 1991 )
Ultraviolet-Sensitive cones in the Goldfish.
Vision Res., Vol.31, No.3, pp.349-352
Frishch, K. V. ( 1925 )
Farbensinn der Fische und Duplizitatstheorie.
Zeit. vergl. Physiol., 2, pp.393-452
袋谷賢吉( 1991 )
ヒラメの色覚の網膜内神経機構
電子情報通信学会論文誌
Granit, R. and G. Svaetichin ( 1939 )
Principles and technique of the electrophysiological analysis
of colour reception with the aid of microelectrodes.
Upsala Lakaref. Forh., 65, pp.161-177
Hamburger, V.( 1926 )
Versuche uber Komplementalfarben bei Elritzen.
Zet. vergl. Physiol., 4, pp.286-304
伊奈和夫・領木快一・東 久美( 1979 )
マダイの色の対する反応性
日水誌, 45, pp.1-5
井上 実( 1978 )
魚の行動と漁法
恒星社厚生閣, pp.66-67
柿元 晧・大久保久直( 1985 )
新潟沿岸域における人工漁礁の総合的研究と事業
新潟県水試, pp.1-327
柿元 晧・大貝政治・池田 至・野田幹雄( 1995 )
魚類の行動学的研究
水産大学校増殖学科研究報告書, pp.1-11
Kawamura, G., W. Nishimura., S. Ueda., and Nishi, T. ( 1981 )
Color Vision and Spectral Sensitivity in Tunas and Marlins.
Bulletin of the Japanese Society of Scientific Fisheries, 47 ( 4 ), pp.481-485
Kawamura, G. ( 1981 )
Recording of C-type S potential from the Retinae of Sparidae.
Bulletin of the Japanese Society of Scientific Fisheries, 47 ( 6 ), p.825
川村軍蔵・法花正志・荒牧孝行・新村 巌( 1991 )
マダイによるHeNeレーザー光線視認
日水誌, 57 ( 3 ), pp.421-424
川本信之( 1959 )
魚類生理生態学, 第2編漁撈生理生態,
第11章感覚及びその機構,2.視覚, 1.走光性
恒星社厚生閣, pp.191-194
Kobayashi, H. ( 1962 )
A Comparative Study on Electroretinogram in Fish,
with Special Reference to Ecologocal Aspects.
ChapterⅢ. Spectral sensitivity of fish retina and its ecological aspect
J.Shimonoseki Coll.fish., 11 ( 3 ), pp.407-538
小山長雄( 1978 )
アユの生態
中公新書
桑原万寿太郎( 1983 )
感覚-行動の生物学
桑原万寿太郎,森田弘道編,岩波新書, p.9
MacNichol. E. F. Jr. ( 1964 )
Retinal mechanisms of color vision.
Vision Res., 4, pp.119-133
Marks, W. B. ( 1965 )
Visual pigments of single goldfish cones.
J. Physiol., ( London ), 178, pp.14-32
McCleary and Bernstein ( 1959 )
A unique method for control brightness cues in study of color vision in fish.
Physiol. Zool., 22, pp.284-292
Neumeyer, C, ( 1992 )
Tetrachromatic color vision in goldfish :
evidence from color mixture experiments
Journal Comparative physiology A, 171, pp.639-649
西島りさ子( 1995 )
魚類の視精度に関する組織学的検討
東京水産大学,平成6年度,卒業論文
丹羽宏( 1991 )
魚類生理学, 12.視覚,§2.視覚の情報処理機構
板沢靖男・羽生 功編,恒星社厚生閣, pp423-441
大山 正著( 1994 )
色彩心理学入門
中央公論社, pp.53-79
Tamura, T. and H. Niwa ( 1967 )
Spectral Sensitivity and color vision of fish as indicated by S-potential.
Comp. Biochem. physiol., 22, pp.745-754
田村 保( 1977 )
魚類生理, 20.視覚,§14.色彩視親
川本信之編,恒星社厚生閣, pp.440-444
田村 保( 1977 )
魚類生理学概論, 11章.感覚,§2.視覚, 4.色彩感覚
田村 保編,恒星社厚生閣, pp.237-240
Tomita, T., A. Kaneko, M. Murakami and E. L. Pautler ( 1967 )
Spectral response curves of single cones in the carp.
Vision Res., 7, pp.519-531
張 秀梅・有元貴文( 1993 )
漁獲技術のための魚類の視覚研究序説
水産の研究, 12巻, 4号( 65 ), pp.35-43


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}