学位論文内容要旨
【目的】魚類の視覚機能として,運動視は形態視とともに個体の生存の上で重要であり,餌をとり,あるいは危険を回避するといったさまざまな行動において,運動物体の認知あるいは識別の能力が大きな役割を果たしている。この運動視の特性について,マダイPagrus majorとコイCyprinus carpioを実験魚としてストロボ光による網膜活動電位を測定し,臨界融合頻度に及ぼす刺激光条件と水温の影響について検討した。
【方法】実験魚としてマダイとコイを使用し,網膜活動電位の測定を行った。実験魚を300ppmのFA100溶液で麻酔し,これをシールドボックス内に静置した状態で100ppmの薄いFA100溶液で灌流しながら実験に供した。ステンレス鋼線で作製した関電極を角膜を通して硝子体に差し込み,不関電極を眼球の後部に挿入し,網膜活動電位を導出した。刺激光源としてはストロボスコープ(発光強度170 lx-sec,単発閃光時間20~22μsec) を用い,光強度及び照射頻度の条件を変化させて網膜電位図(ERG)を記録した。本実験では,ストロボの連続閃光刺激に対してERGの振幅がゼロに近づき,波形が平坦になった段階の閃光頻度をERG臨界融合頻度とした。実験項目は以下の4通りで実施した。1)暗順応状態のマダイを用い、光強度による網膜電位の振幅の変化を測定した。光強度は透過率の異なるフィルターを用いて1.25,2.5,5,10,12.5,25,50,100%の8段階に設定した。2)断続的なストロボ光照射による網膜電位の振幅の経時変化について,マダイを実験魚として明順応・暗順応状態別に調べた。 3)明・暗順応状態のマダイをそれぞれ用いて,等強度のストロボ光頻度を低頻度から高頻度,あるいは高頻度から低頻度で照射し,網膜電位の振幅の変化を比較した。4) 明順応状態のマダイとコイを用いて,10,20,25℃の3段階の水温条件に24時間馴致させ,ERG臨界融合頻度に及ぼす水温の影響を調べた。照射頻度は10~50Hzの範囲とし,低頻度から高頻度の順序で照射した。
【結果】ストロボ光の単発照射について,マダイの網膜電位の振幅は光強度の増加に伴って増大した。また網膜の明・暗順応過程におけるマダイの網膜電位について,明順応過程において振幅の増大が,暗順応過程において振幅の減少が見られた。ERG臨界融合頻度を調べるための実験としては,網膜の順応状態とストロボ光頻度の照射順序が結果に影響することが考えられる。これについて,明順応状態で全体的に網膜電位の振幅は大きく,また低頻度から高頻度という順序で照射した場合に照射頻度の影響を特徴的に示すことが認められた。最後に水温の影響についての実験より,マダイのERG臨界融合頻度は10,20℃ではともに26Hzであったのに対して,25℃で35Hzと高温条件での運動視の向上が確認された。コイの場合には,10℃で29Hz,20℃で32Hz,25℃で37Hzという結果が得られた。
PDFファイルへのリンク
緒言
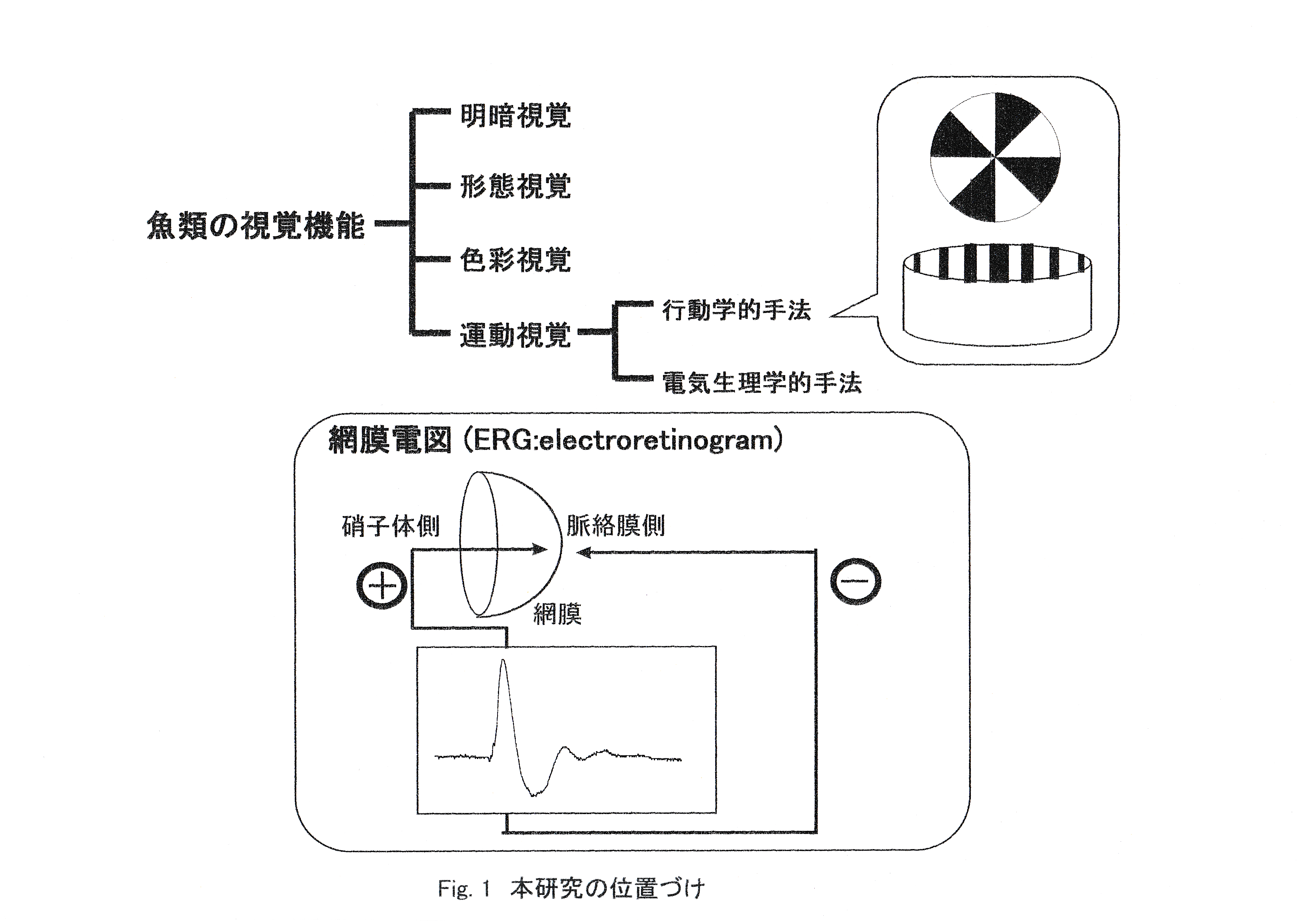
魚類の視覚機能の内でも運動視(Motor vision)は形態視(Form vision)とともに個体の生存の上で重要であり,餌をとり,あるいは外敵を回避するといったさまざまな行動において運動物体の認知あるいは識別の能力が大きな役割を果たしている(張,1993)。 本研究の位置づけをFig.1に示す。 魚類の視覚機能は明るさを感じる明暗視覚,ものの形を見分ける形態視覚,色を見分ける色彩視覚,動くものを眼でとらえる運動視覚に分類される。運動視覚の研究手法は学習や視運動反応を用いた行動学的手法と電気生理学的手法に大別される。
行動学的手法では白黒の回転板を頻度を上げて回転させて,いつ灰色に見え始めるかで臨界融合頻度を求めたり,縞模様をかいた円形の視覚スクリーンを徐々にはやく回転させ,中の実験魚がいつ追従しなくなるかで臨界融合頻度を求める。Beniuc(1933)はトウギョを用いて学習法により実験し,この魚のcffはcff=55/secとしている。すなわち黒白の扇形に染めた円板をゆっくり回転したものに対し-に学習し,一方これを速やかに回転したと同価の灰色に対し+に学習しておく。学習が完成した魚を用い扇形に染めた円板をどの程度の早さで回転した時に灰色円板に対すると同じ反応を示すか検し,cff=55/secを得たのである。Sun-fish(Lepomis,ブルーギル等)やメダカをガラスの円形水槽に入れ,この外側で縞模様を回転させると,魚はこの縞の回転を追う性質がある(menotaxis,保留走性の一種で,optomotor reaction視運動反応ともいわれる。)。この場合縞の回転速度をだんだん上げていくと魚の眼に縞が融合してしまい灰色に見える速度があるはずである。この時には魚はもはや走性を示さなくなる。これによってcffを求めることができる。Wolf and Zerrhan-Wolf(1936)はこの方法を用い明るさをかえて実験し十分明るければcffは50/secと見ることができることを見た(田村,1970)。
電気生理学的手法では刺激光照射に対する網膜電図を記録し,照射頻度を上げた場合にいつ反応がなくなるかで臨界融合頻度を求める。
ここでまず網膜活動電位について簡単に述べることとする。眼の中を眼底から角膜の方に流れる電流があり,これは網膜の静止電流である。網膜に光刺激を与えると過分極が生じ,細胞内の静止電位が一旦負になる。この受容電位は双極細胞,水平細胞,神経節細胞などを介して中枢に伝わっていく。網膜を光で刺激すると,このような多くの神経細胞が活動し,この時の電位変化が網膜活動電位である。この際,網膜をはさんで硝子体側と脈絡膜側とに電極をおき,電位変化を描画記録したものが網膜電図(Electroretinogram ERG)と定義される(張,1992)。
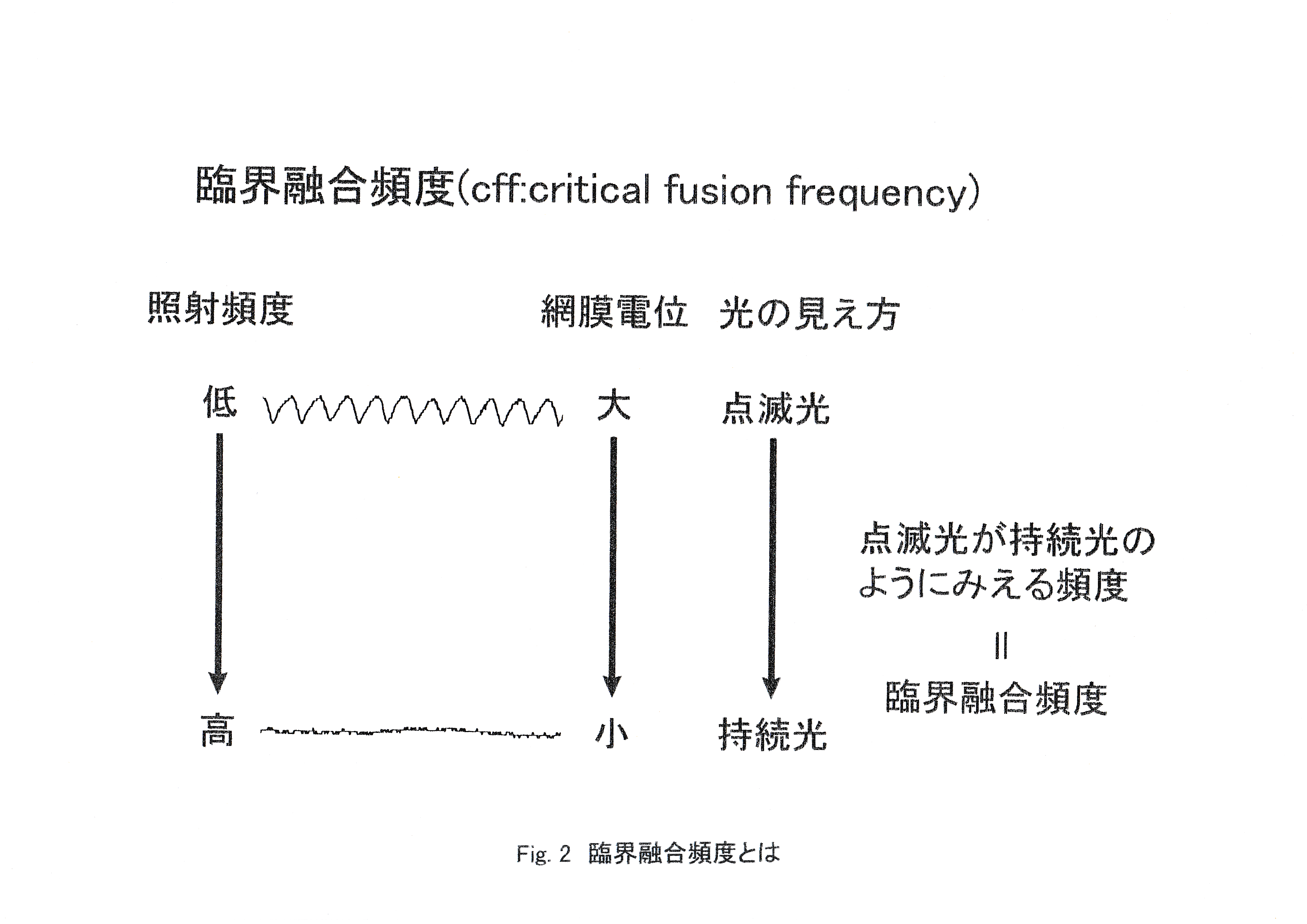
運動視の指標として臨界融合頻度があげられる。Fig.2に説明の図を示す。 ストロボ光の照射頻度を低頻度で照射すると,1つ1つの光に対応して網膜電位の波形が確認でき,これを高い頻度で照射すると,その振幅は小さくなっていく。光の見え方は照射頻度が低いときは点滅光として認識できるが,途中から持続光のように見える。
臨界融合頻度は点滅している光の頻度がある一定値以上になると,点滅が融合して持続光のように見える頻度であり,チラツキ光が連続して見える臨界値を融合頻度とし,この値が高いとき視覚の時間的分解能が優れていることを意味する(宗宮・丹羽,1991)。本研究ではストロボの連続閃光刺激に対してERGの振幅がゼロに近づき,波形が平坦になった段階の閃光頻度をERG臨界融合頻度(CFF)と定義した。
本研究では運動視の特性について,マダイPagrus majorとコイCyprinus carpioを実験魚としてストロボ光による網膜活動電位を測定し,臨界融合頻度に及ぼす刺激光条件と水温の影響について検討した。
始めに魚類のERGに関する基礎的検討としてマダイを用いて,単発照射時の光強度による網膜電位,網膜電位の経時変化,連続照射時のERG波形と照射頻度の影響を調べた。 この結果を踏まえて,次に魚類の運動視覚に関する基礎的検討として,マダイのERG臨界融合頻度,コイのERG臨界融合頻度における水温条件の影響について調べた。
第1章 魚類のERGに関する基礎的検討
第1節 マダイの光強度ごとの網膜電位
緒言
本章では,魚類の視覚機能を解明するために,電気生理学的手法を用い,特に光強度,発光頻度,水温などの条件による網膜の応答に関して,網膜活動電位を指標として検討した。マダイを実験魚として用い,ERGの測定により刺激光強度,網膜の明暗順応過程における網膜感度の変化,順応状態と照射手順によるERG臨界融合頻度の変化などを測定し,電気生理学的手法を用いた基礎実験を行った。
本章ではマダイの網膜感度の基本的特性を明らかにする。第一節として刺激光強度によるERGの変化を調べる。
方法
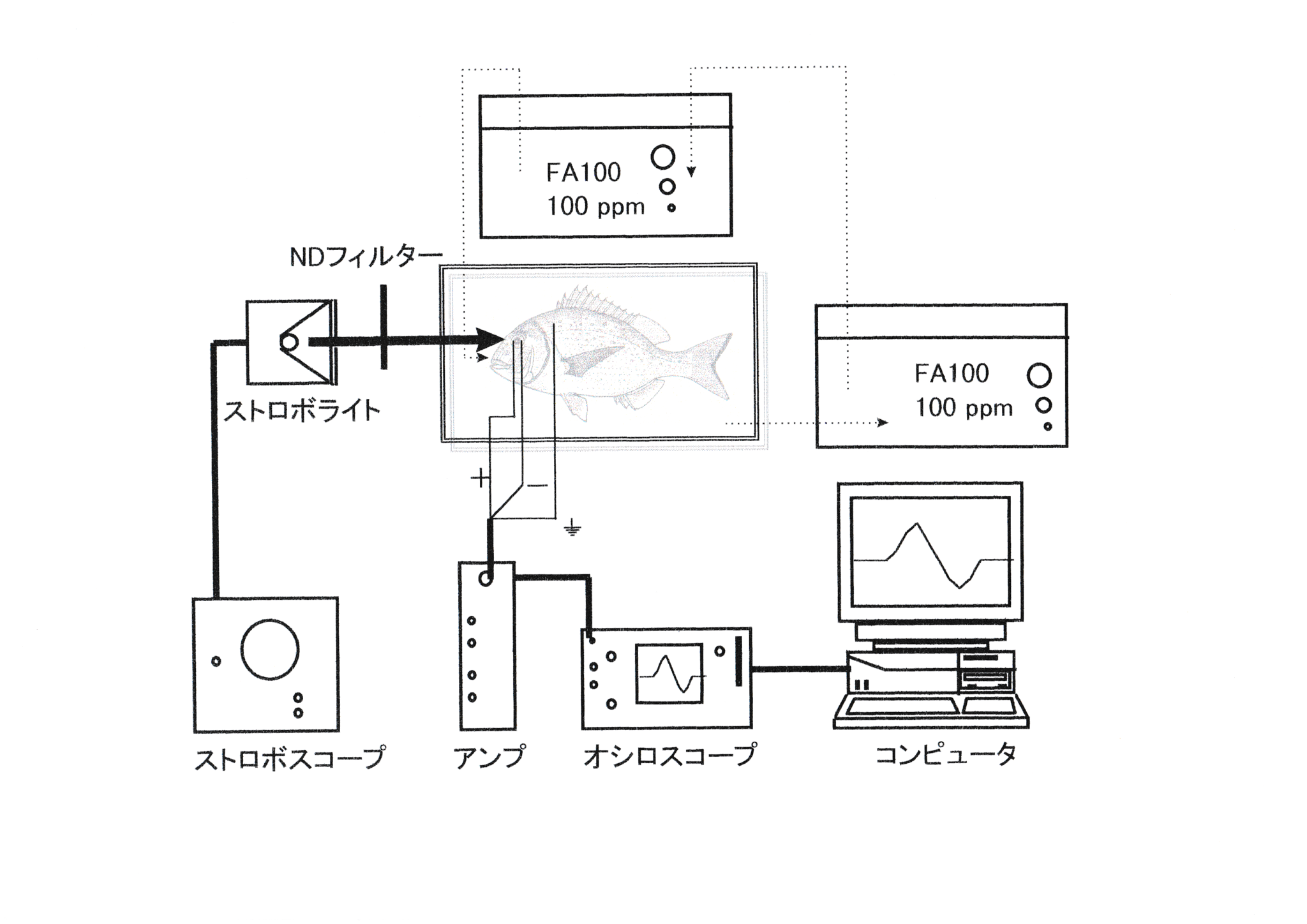
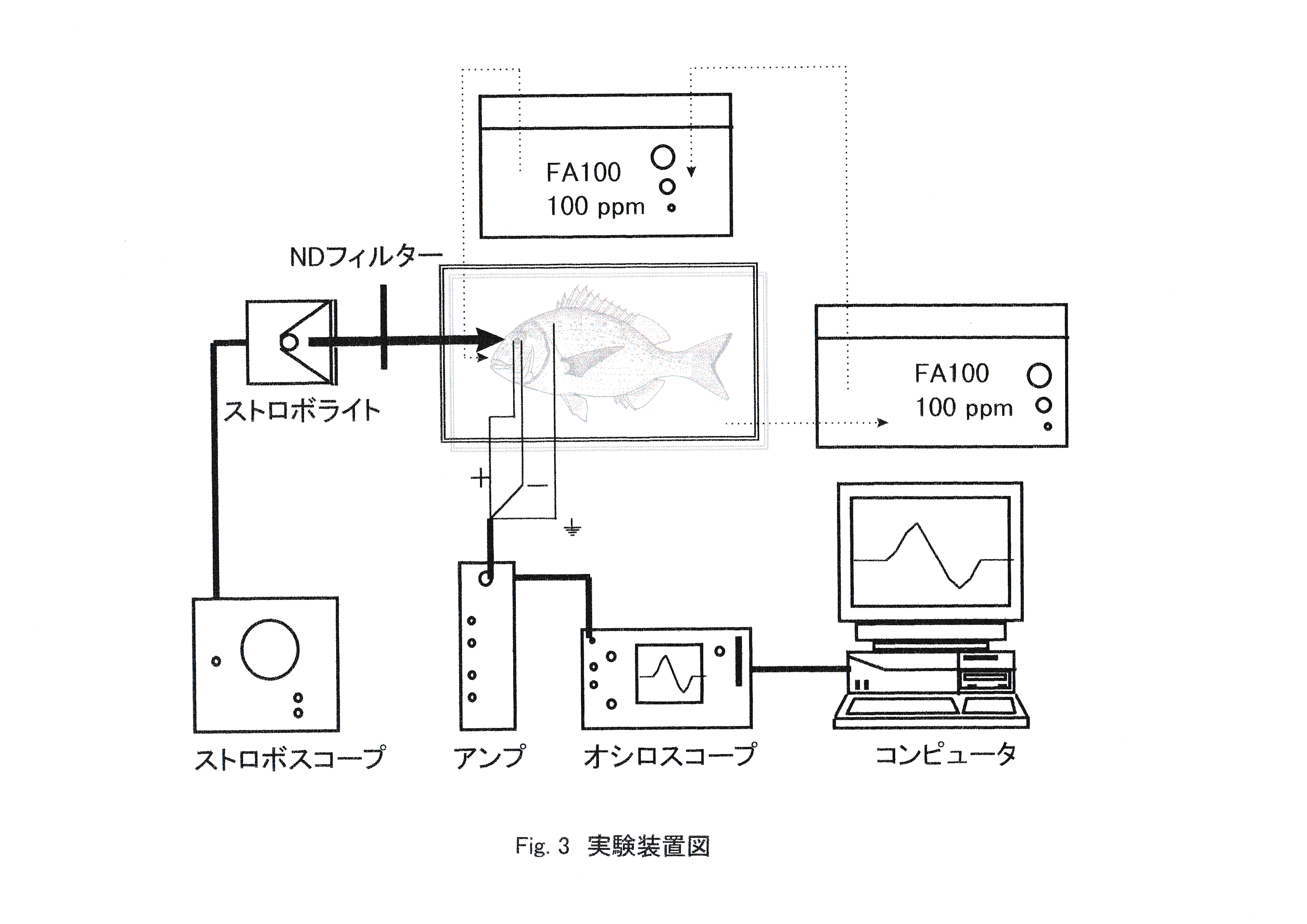
実験魚として体長21.2 cmのマダイ(神奈川栽培センターで飼育された2年魚を平成8年5月31日より東京水産大学魚群行動学研究室飼育室内で飼育した)を一尾使用した。実験は平成8年12月13日から14日にかけて行った。Fig.3に実験装置図を示す。 実験魚は13日の17:00から暗順応させた。14日の3:30に実験魚を3/10000(300ppm)のFA100(田辺製薬株式会社製,魚類・甲殻類麻酔剤)溶液で麻酔し,筋弛緩剤(塩化ツボクラリン)0.02mlで不動化した。実験魚は暗幕内のシールドボックスに静置した状態で1/10000(100ppm)の薄いFA100溶液で灌流しながら実験に供した。水温は10℃一定とした。作業の間は間接照明として直流光源のみを使用した。ステンレス鋼線(エム・ティ技研社製,0.4φ×200m/mコート付)で作製した関電極を角膜を通してガラス体に差し込み,不関電極を眼球の後部に挿入し,導出された網膜活動電位を高感度ACアンプ(日本光電,AB-632J)で増幅し,デジタルストレージスコープDIGITAL STORAGESCOPE(岩通DS6612C,DS8606C)に表記すると同時にICメモリカード(128KB)に記録した。照度計{デジタル照度計(MINOLTA,T-1M)}を設置することによりストロボ光の照射と波形の発生との一致を確認した。 記録された波形のデータをフロッピーディスク(1.2MB)に転送し,コンピュータ(エプソンPC-286LS,NEC PC-9821V13)により解析を行った。アンプの感度設定は0.05mV/div,高域遮断周波数は300Hzとし,オシロスコープの時定数は2secに設定した。刺激光としてストロボスコープ(菅原研究所,PS240型)を用い,単発照射した。光強度は8-12μで一定とした。閃光周波数は80~26500rpm(1.4~440Hz)の範囲で,実験条件として発光強度は170 lx-sec,閃光頻度2~50Hzの範囲内で測定を行った。ストロボライトは暗幕内で実験魚との距離が40cmとなるように設置した。ストロボライト発光部の面積は3.14cm2とした。NDフィルター{東芝硝子株式会社製色ガラスフィルター(Neutral-density filter)ND-10,ND-25,ND-50}を用いて,光強度を0 (control),1.25,2.5,5,10,12.5,25,50,100%の9段階に設定し,15分おきにこの9段階の刺激光に対する網膜の反応をERGとして記録し,網膜電位の振幅を測定した。
結果
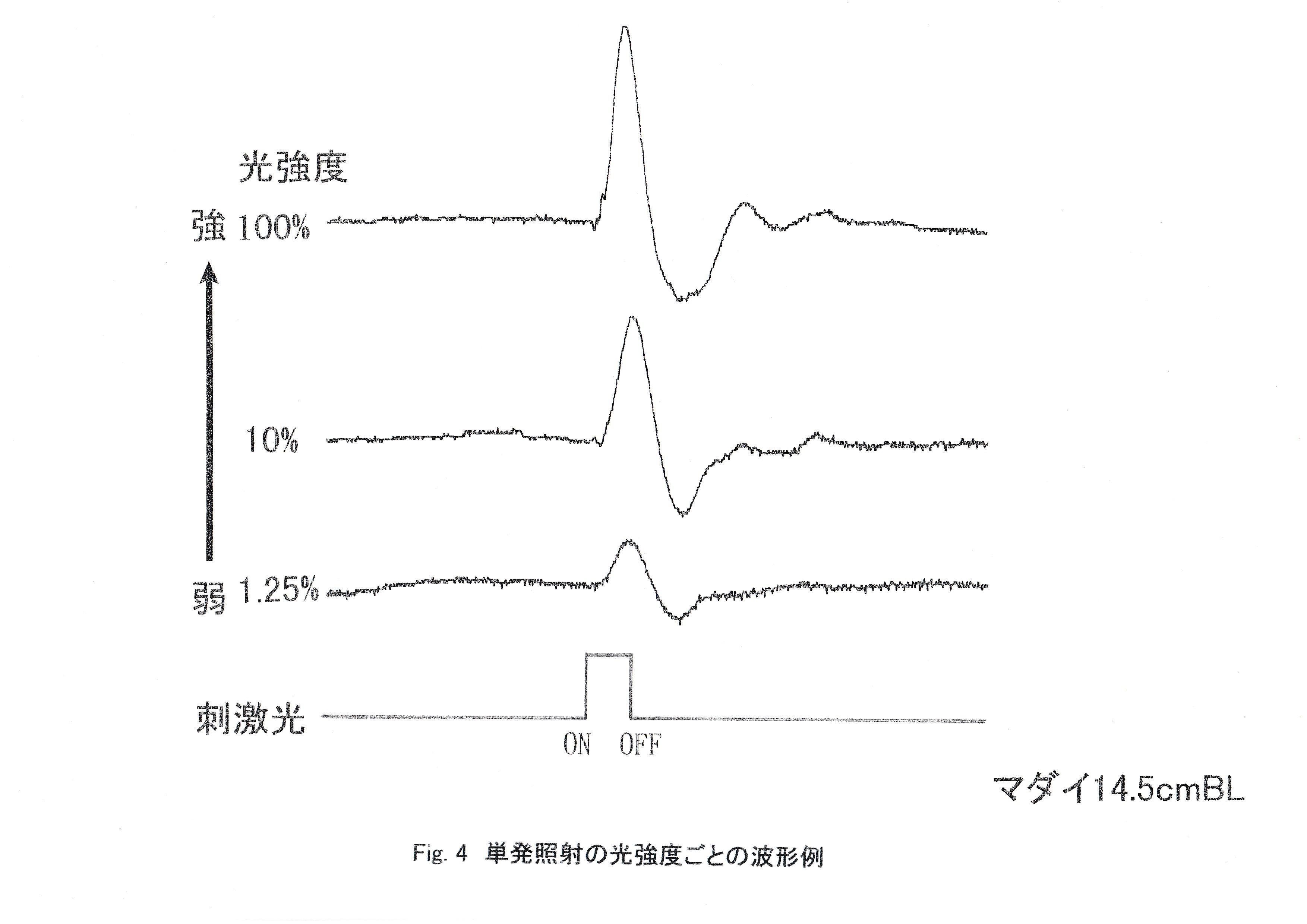
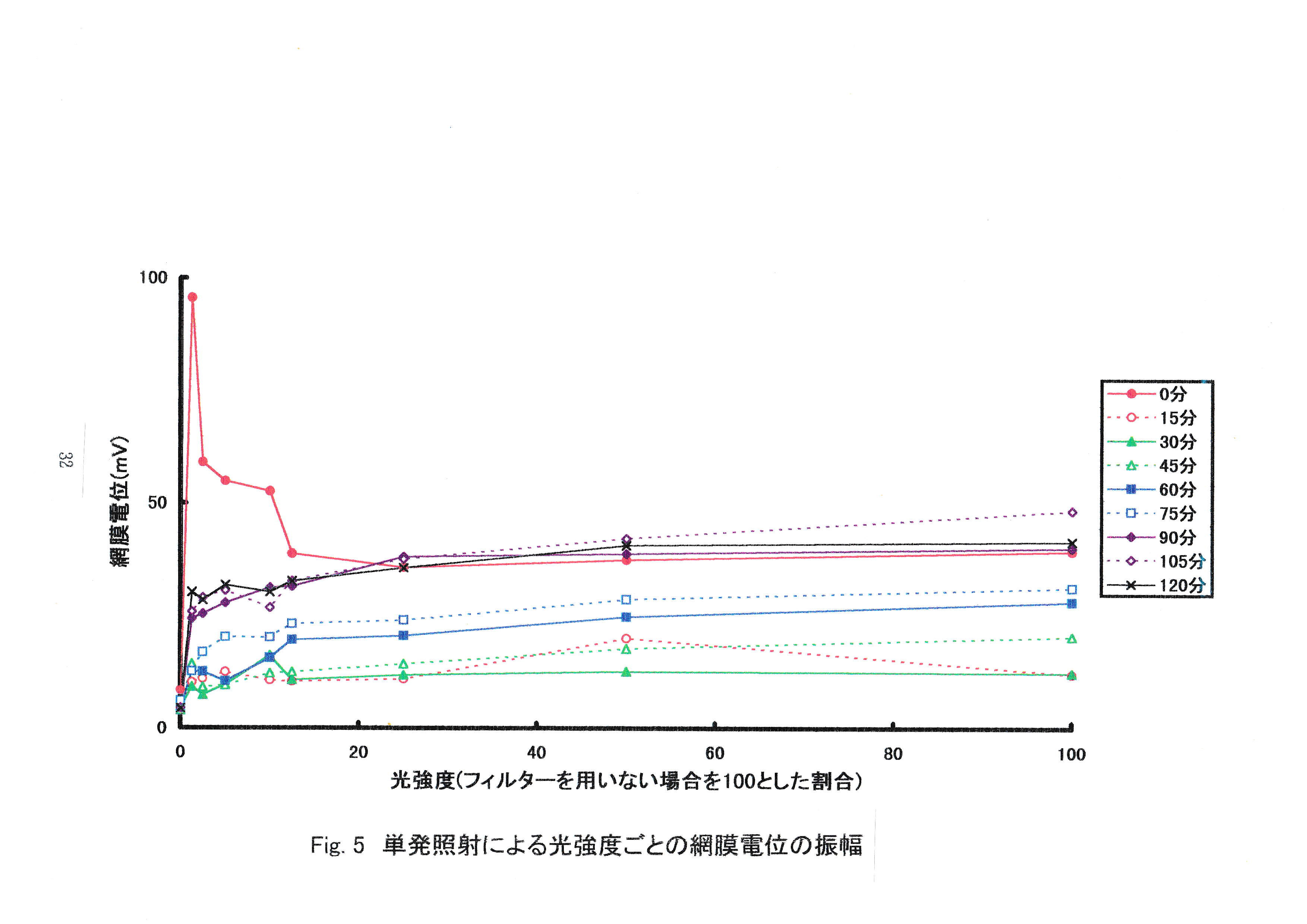
Fig.4にストロボ光単発照射の光強度ごとの波形例を示す。 上の3つの波形がERGで,一番下の線が刺激光のON-OFFを示す。ONで点灯と同時に波形が現われ,OFFの消灯で波形が完了している。刺激光強度が100,10,1.25 %と弱くなるにつれて,ERGの振幅が小さくなっている。Fig.5にストロボ光単発照射による光強度ごとのマダイの網膜電位の振幅を示す。 最初の照射5回までは光強度の増加に伴う網膜電位の振幅の減少がみられた。
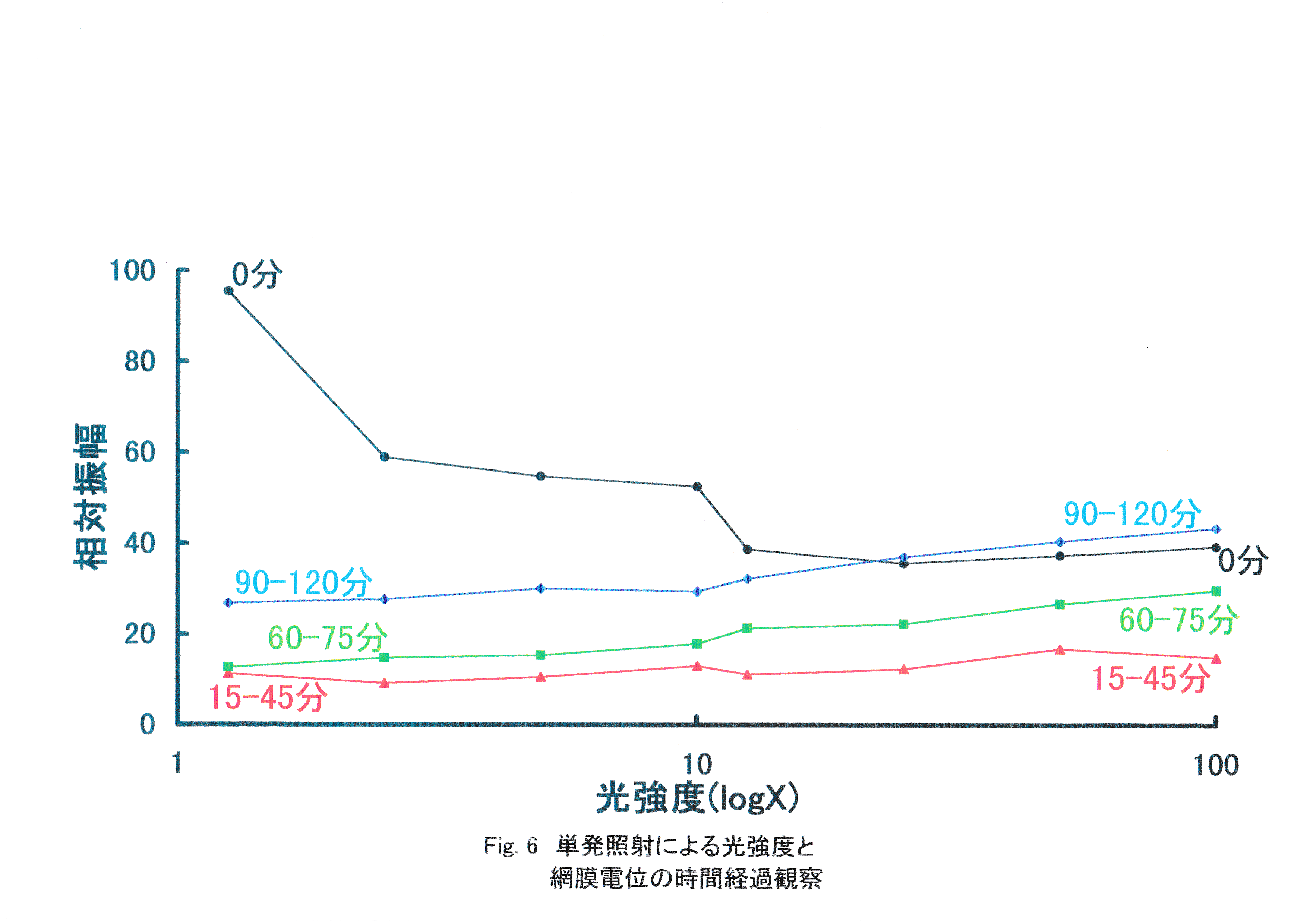
ストロボ光照射開始0分後と15,30,40分後と60,75分後と90,105,120分後の振幅の平均を求め,光強度を対数で示すと,Fig.6になる。 照射開始から15,30,45分後・60,75分後・90,105,120分後とそれぞれの光が強いときに電位が大きくなり,時間を経過するごとに振幅全体は大きくなり,暗順応に応じて感度が高くなっていく様子が確認できる。
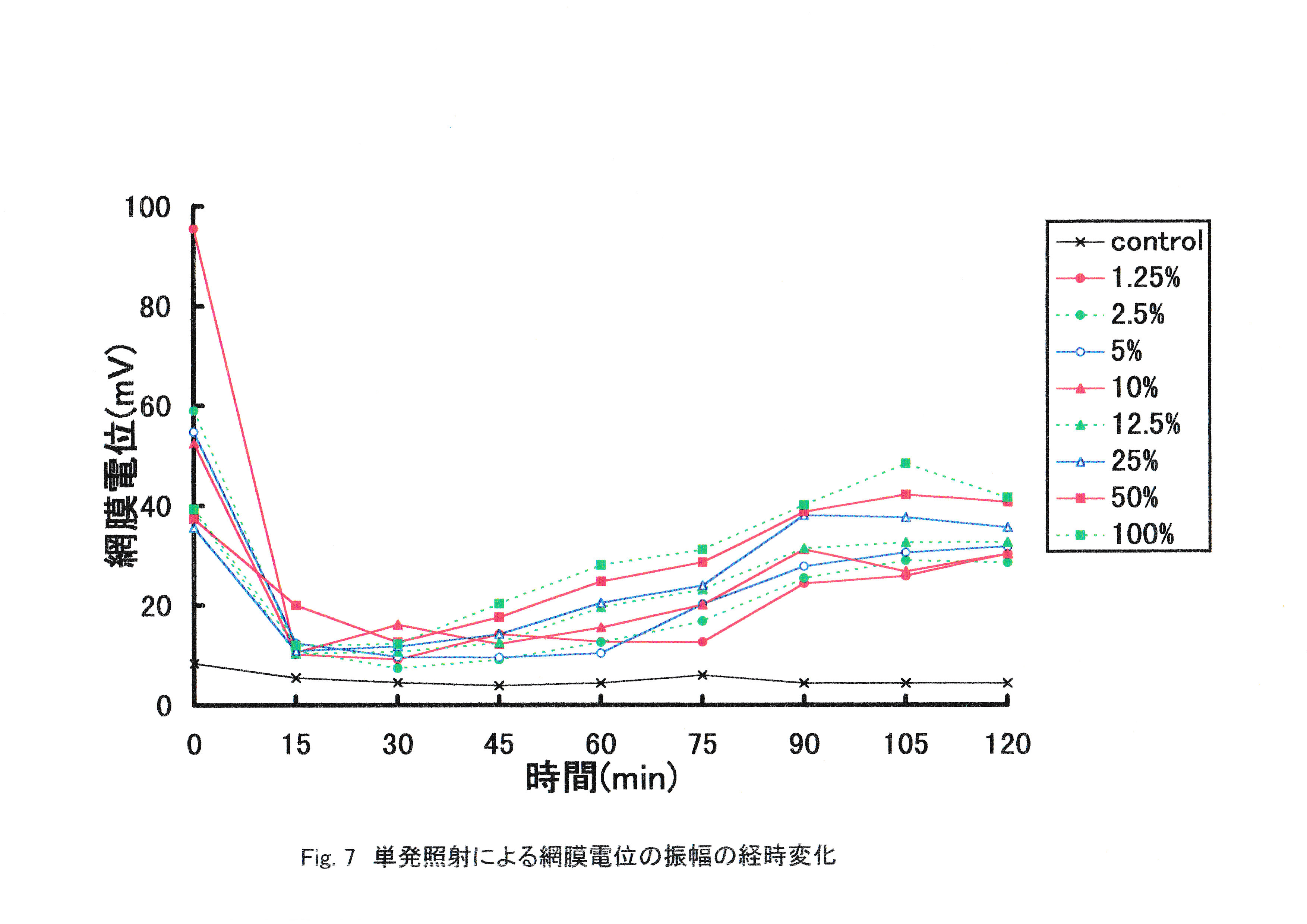
Fig.7にストロボ光単発照射によるマダイの網膜電位の振幅の経時変化を示す。 照射開始時の網膜の反応が最も大きく,15分経過で網膜電位の振幅は最も小さくなり,その後振幅が増大する結果となった。
考察
最初の照射5回まで光強度の増加に伴う網膜電位の振幅の減少がみられたのは,最初の照射5回までで完全な暗順応状態となっていた網膜が光エネルギーを受けることで慣れたことによると考えられる。最初の照射5回に対する網膜の振幅の減少は照射された光強度が同光量であればさらに落ち込むと予想される。照射開始から15,30,40分後・60,75分後・90,105,120分後と時間を経過するごとに網膜の反応が大きくなり,暗順応していたと考えられる。照射開始15分で暗順応から明順応への中間状態,その後中間状態から暗順応状態となり,時間経過を追うごとに振幅が増大する傾向になったと考えられる。それぞれの順応状態で光強度が強ければ網膜の反応は大きくなった(感度が高くなった)と考えられる。
第2節 マダイの順応状態別の網膜電位の経時変化
緒言
光強度の変化に対応する眼の調節作用は順応といわれ,明所から暗所に移ったときの光に対する視感度の変化過程を明順応(Dark adaptation),暗所から明所に移行したときを明順応(Light adaptation)と定義される(中村,1969)。
本節では網膜の明・暗順応過程における網膜感度の変化を調べた。
方法
実験は平成8年11月13-15日に行った。実験装置および測定条件の設定は前節と同様である。実験魚としてマダイを用いた。体長18cmのマダイを13日の18:00から24:00まで暗順応させた。体長18.2cmのマダイを15日の9:00から18:20まで明順応させた。順応させた後に麻酔して暗幕内に搬入し,ストロボ光を断続的に実験魚の網膜に照射した。ストロボ光の照射頻度は2Hz(120rpm)で一定とした。ストロボ光照射開始から59分2秒の間に60回,1分毎に2秒間のERGを記録し,2秒間で得られた4つの値の平均値を1分毎のERGとした。
結果
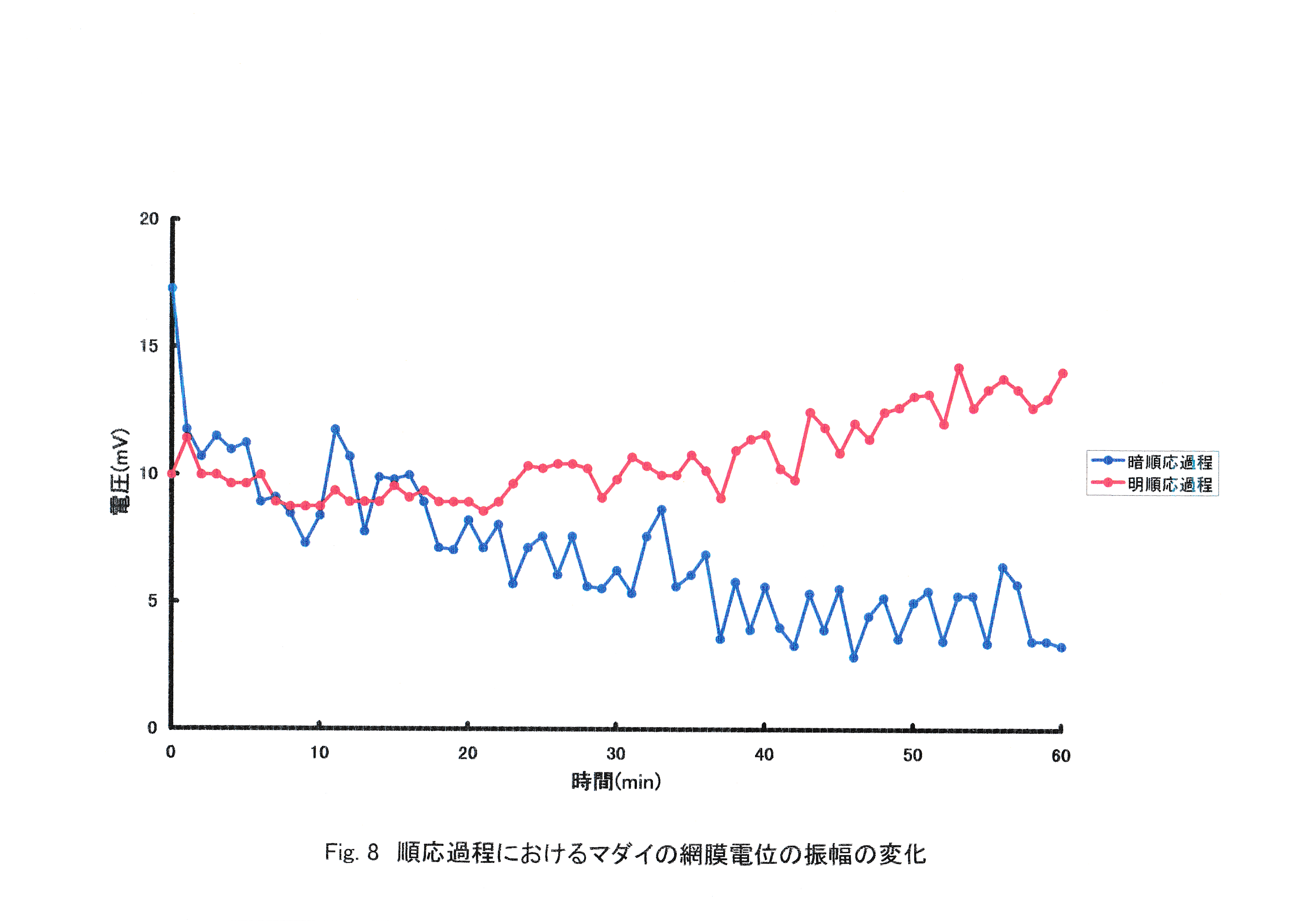
Fig.8にERGの振幅の経時変化を示す。 明順応させた実験魚は時間を追うごとに網膜電位の振幅が減少する傾向がみられた。暗順応させた実験魚では最初の20分間は網膜電位の振幅は減少し,それ以降は振幅が増大する傾向がみられた。
考察
暗順応させた実験魚で振幅の増大が見られたことから,光照射による明順応が生じていないと考えられる。明順応させた実験魚で振幅の減少が見られたことから,暗順応過程で感度が増加する手前だったと考えられる。
網膜の順応状態が暗から明,明から暗順応に移行する中間状態であったと考えられる。刺激光照射中は明順応・暗順応とも同じ実験設定であるのでこの実験を継続して行っていたならば網膜電位の振幅の増減に差はなくなるはずである。
実験設定において実験魚を明順応させるには背景光を利用し,暗順応させるには刺激光を照射する間隔をもっとあける必要があったと思われる。
第3節 ストロボ光照射開始時・照射中・照射終了時の網膜電位
緒言
ストロボ光を持続的に照射した場合,ストロボ光照射中の網膜電位はもちろん,ストロボ光照射開始時・終了時の網膜電位も記録される。ストロボ光照射頻度を変えた場合,ストロボ光照射開始時・照射中・終了時の網膜の反応がどのように変化するかを観察した。
方法
実験は平成10年12月30日に行った。実験装置および測定条件の設定は前節と同様である。体長14.5cmのマダイ1尾を用いた。実験魚は照度1020 lxの環境下で13:00~16:20まで3時間20分明順応させた。水温は25℃とした。ストロボ光照射頻度は10,20,30,40,50Hzの5通りとした。ストロボ光照射開始時・照射中・終了時の網膜電位を記録した。
結果
Fig.9にストロボ光ON-OFF時の波形例を示す。 ストロボ光の照射開始に対する大きな波形,照射中の点滅1回ずつに対応する波形,照射終了を示す波形の3種類の波形が観察された。
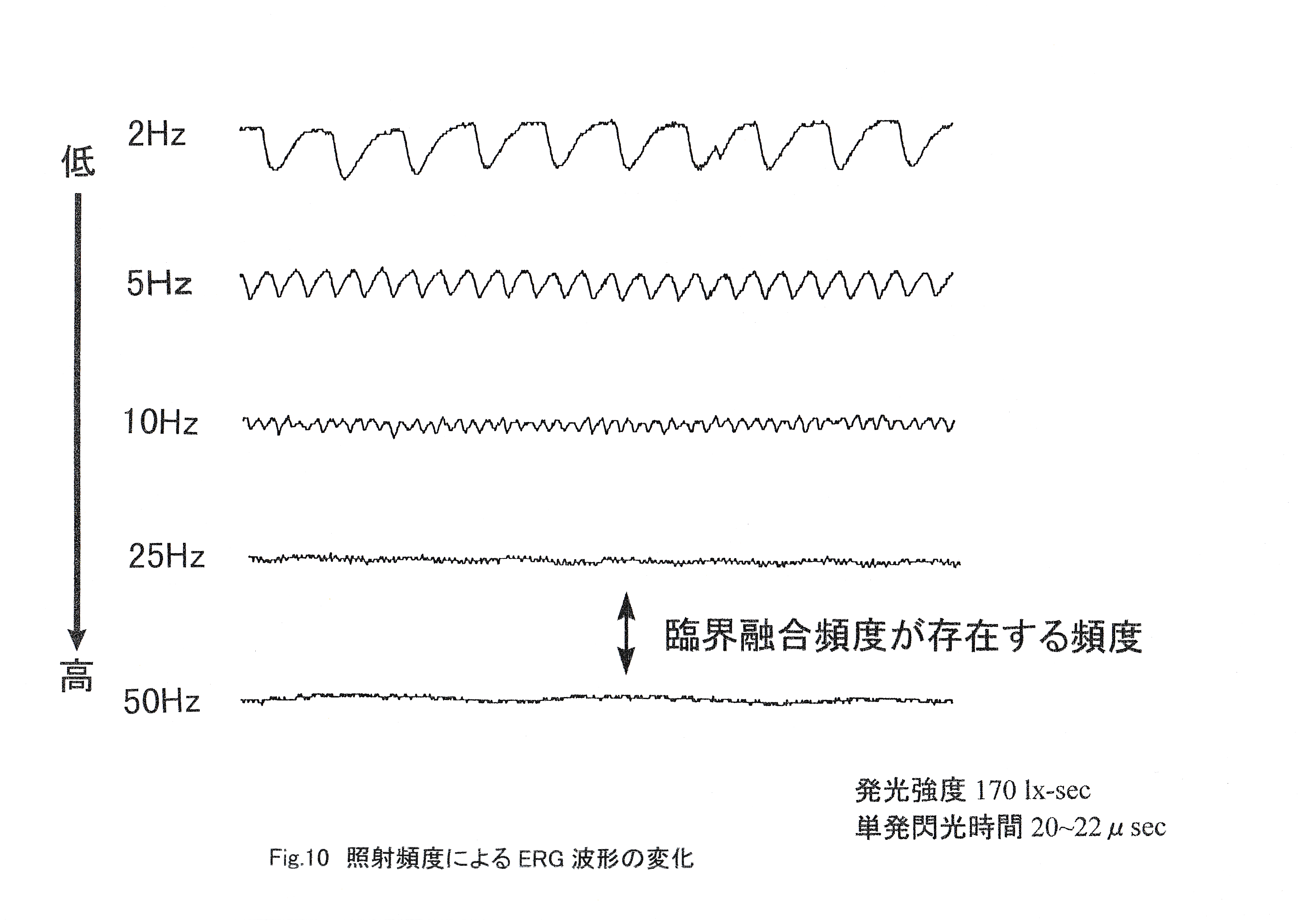
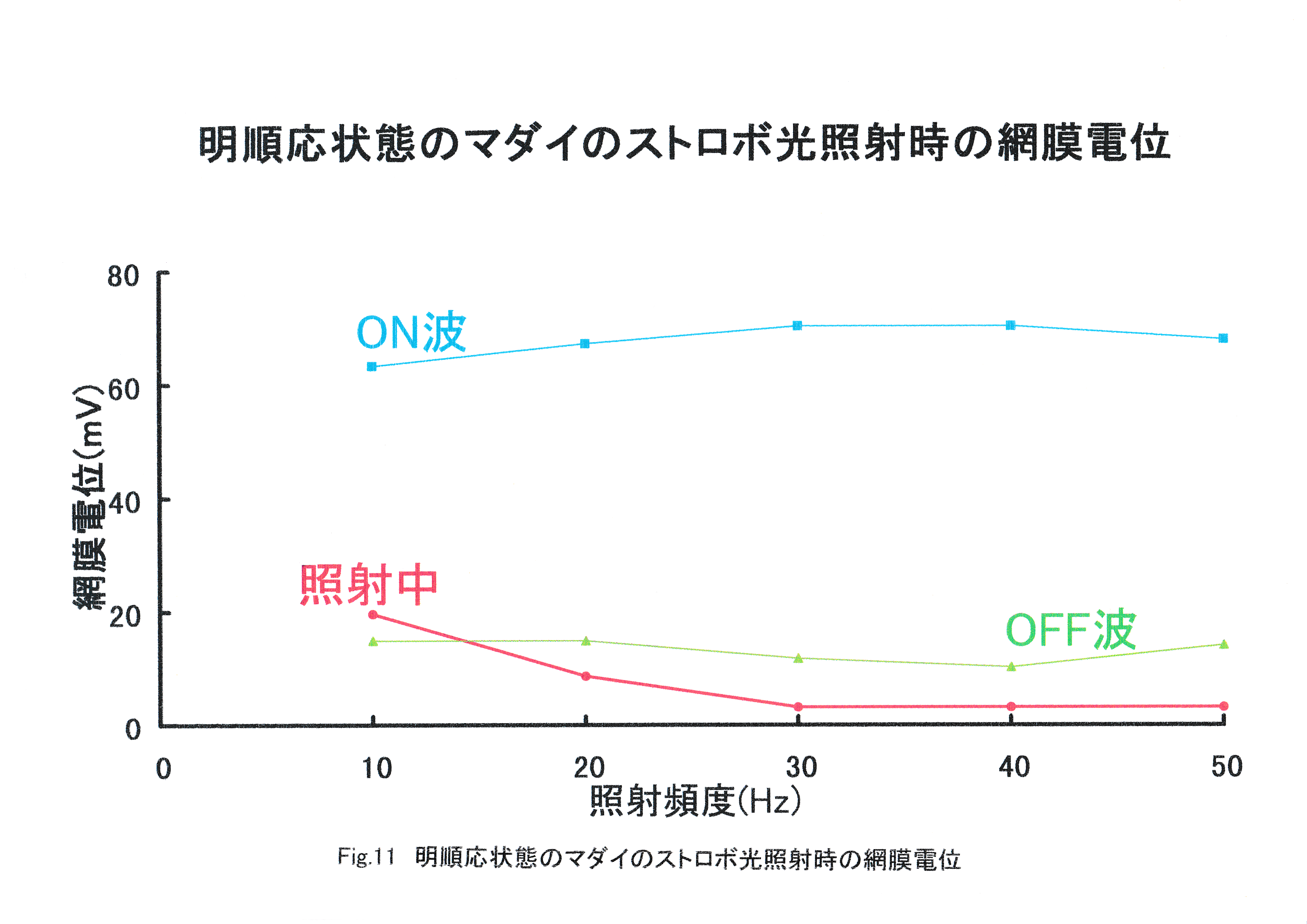
Fig.10に照射頻度ごとのストロボ光照射中のERGの例を示す。 照射頻度が高くなると振幅が減少しているのがわかる。25Hzでは僅かに一つずつの波形が確認できるが,50Hzでは基線のノイズに重なっており,25Hzから50Hzの間に臨界融合頻度があることがわかる。Fig.11に照射頻度ごとのストロボ光照射開始時・終了時の網膜電位の振幅とストロボ光照射中の網膜電位の振幅を示す。 照射頻度の増加に伴い,ストロボ光照射中の網膜電位の振幅は減少する傾向がみられた。ストロボ光照射開始時・終了時の網膜電位の振幅にはストロボ光の照射頻度の増加に伴う変化は見られなかった。
考察
照射頻度が低いときはストロボ光の点滅一回に対してON-OFF光に対する反応と同様の反応がみられるが,照射頻度が高まると点滅一回に対する反応ではなく,ストロボ光照射時全体をON-OFF光照射時とする反応に移行していくものと考えられる。照射頻度の増加に伴い,ストロボ光照射中の網膜電位の振幅が減少する傾向がみられたことから,ストロボ光照射中の網膜電位の振幅がストロボ光の照射頻度に対する反応を示すものと推察された。
第4節 順応状態と照射頻度がマダイの網膜電位に及ぼす影響
緒言
本節ではマダイの網膜の順応状態が明順応の場合と暗順応の場合のストロボ光に対する網膜電位の振幅の大きさを比較する。また,ストロボ光を低頻度から高頻度に照射する場合と高頻度から低頻度に照射する場合で網膜電位の振幅の大きさを比較する。これにより,マダイのERG臨界融合頻度を求めるにはどのような手順と網膜の順応状態の設定が必要であるかを調べる。
方法
実験は平成9年2月5日から4月28日にかけて行った。実験装置および測定条件の設定は前節と同様である。体長18-20cmのマダイを5時間以上明順応させたもの2尾と5時間以上暗順応させたもの2尾の計4尾を用いた。照射するストロボ光の頻度は10,20,30,40,50Hzを設定し、低頻度から高頻度に照射する場合と高頻度から低頻度に照射する場合を設定した。ストロボ光は5分間置きに照射し,明・暗順応状態別に照射頻度10-50Hzを10Hzずつ順次に上げた場合と,下げた場合で記録した。
結果
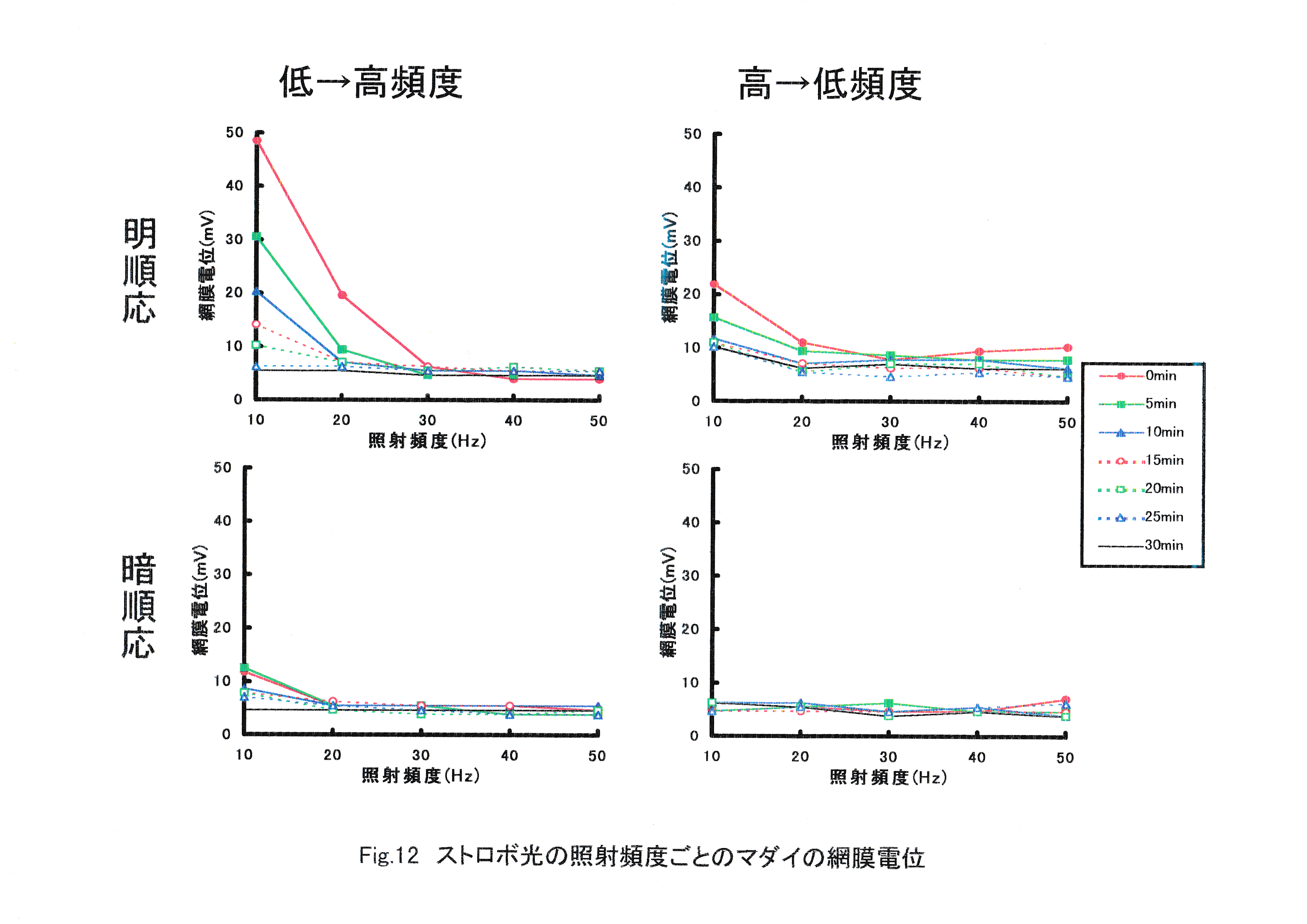
Fig.12にストロボ光照射頻度ごとのマダイの網膜電位を示す。 照射頻度の増加に伴う網膜電位の振幅の減少が見られた。明順応させたマダイの網膜電位の振幅は暗順応させたマダイの振幅よりも大きかった。低頻度から高頻度に照射する場合と高頻度から低頻度に照射する場合では、低頻度から高頻度に照射する場合の方が反応が大きかった。網膜電位の振幅は網膜が明順応状態で低頻度から高頻度に照射する場合に最大値を示した。
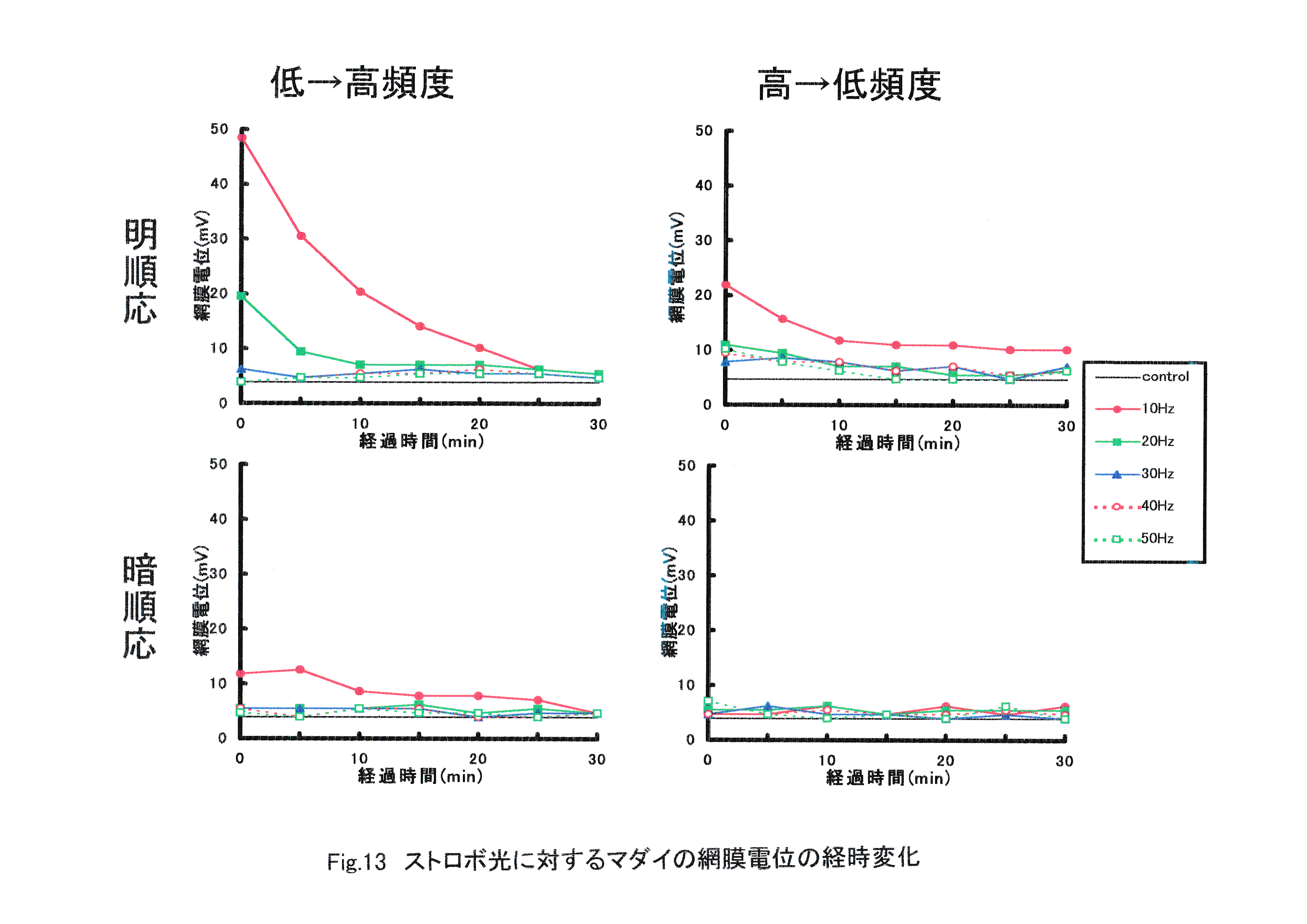
Fig.13にストロボ光に対するマダイの網膜電位の経時変化を示す。 時間の経過していない最初の照射に対する網膜の反応が大きかった。すべての実験魚で照射開始から約30分後には網膜電位の振幅が見られなくなった。
考察
ERG臨界融合頻度は20~40Hzの間に存在すると考えられた。低から高頻度に照射する場合と高から低頻度に照射する場合では、低から高頻度に照射する場合の方が反応が大きくなった。高から低頻度では傾向は同じだが,電位が全体に小さく,照射頻度の影響が見にくくなる。 ERG臨界融合頻度を知るために特徴的な網膜電位を導出するには、明順応させた実験魚を用いて、ストロボ光の照射頻度を低頻度から高頻度に変化させる必要があると考えられた。経過時間による感度の低下は顕著であり,照射頻度を変えて波形変化の違いを見るには短時間内での実験が要求される。
第2章 魚類の運動視覚に関する基礎的検討
第1節 ストロボ光によるマダイのERG臨界融合頻度
緒言
第一章よりストロボ光に対するERGは網膜の光強度,順応状態,照射頻度によって,その振幅に差を生じることが明らかとなった。 ERG臨界融合頻度を求めるには網膜が完全に明順応状態となっている実験魚を用いて,ストロボ光の照射頻度を徐々に上げていく設定で網膜電位を記録する必要があった。本章ではERG臨界融合頻度に及ぼす水温の影響を調べるため,照射頻度ごとのERGを記録した。第一節としてマダイのERG臨界融合頻度について検討した。
方法
実験は平成10年10月14日から12月30日にかけて行った。実験装置および測定条件の設定は前節と同様である。実験魚としてマダイ6尾を用いた。実験魚は一尾ずつ暗幕内に搬入し,クーラーを用いて水槽内で24時間,水温10,20,25℃に馴致させた。灌流水温もこれに対応させて調節した。水温に馴致させると同時に5時間以上かけて実験魚を完全に明順応状態とさせた。照射頻度はまず10,20,30,40,50Hzの5段階に設定し,ストロボ光の照射頻度を10Hzごとに増加させ,臨界融合頻度の存在する照射頻度を10Hzの間隔で求めた。次にその間隔10Hzの照射頻度を1Hzごとに増加させ,ERG臨界融合頻度を決定した。臨界融合頻度は最終的に平衡時の振幅と差がなくなったときの照射頻度と定義した。各水温条件で2個体について記録を行い,実験終了後に生理状態の良かった個体についての結果を用いて解析した。
結果
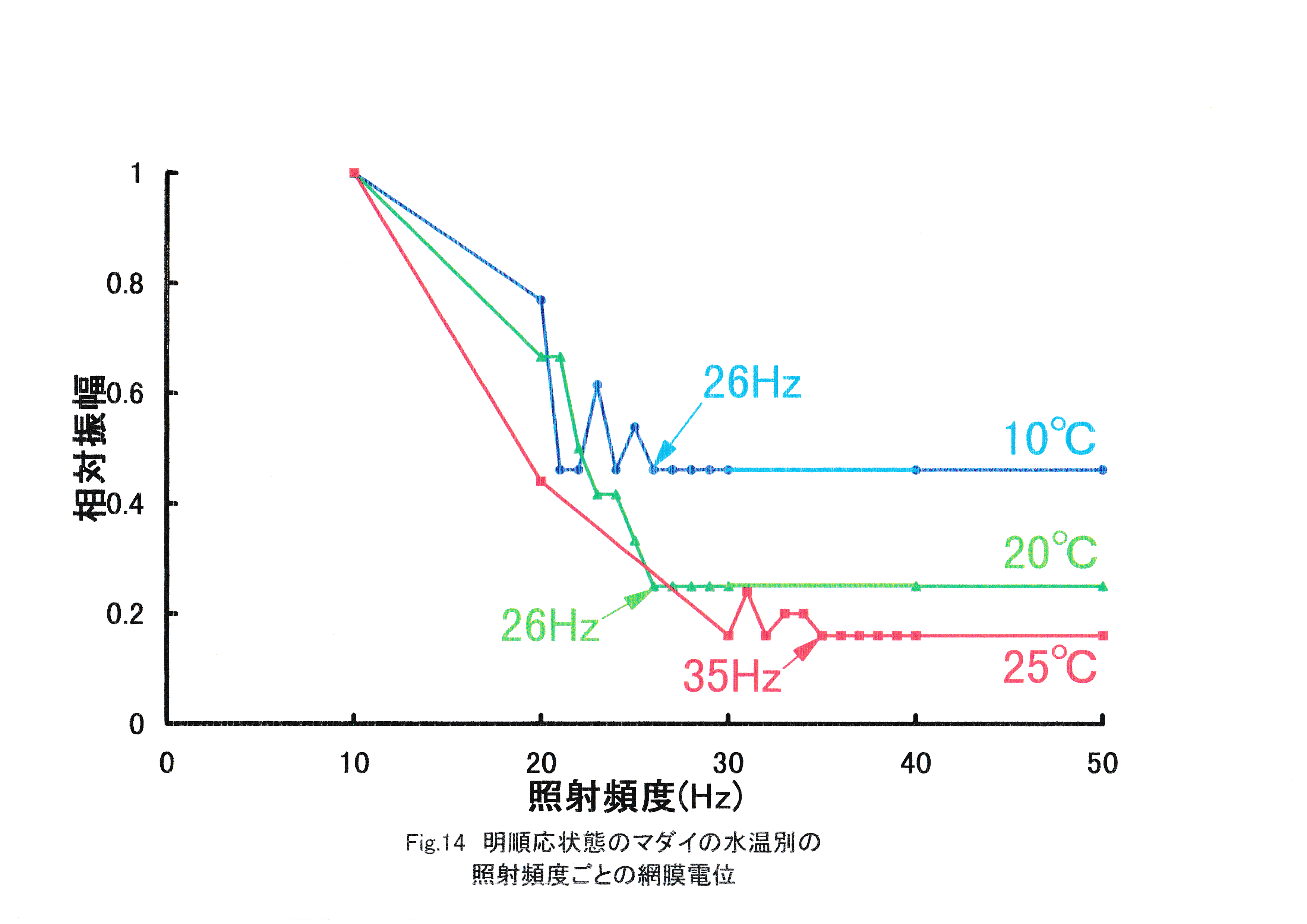
Fig.14に明順応状態のマダイの水温別照射頻度ごとの網膜電位を示す。 全実験魚で照射頻度の増加に伴う網膜電位の振幅の減少が見られた。水温10℃の実験魚のERGは21,22,24,26-50Hzで基線と区別がつかなくなった。水温20℃の実験魚のERGは26-50Hzで基線と区別がつかなくなった。水温25℃の実験魚のERGは30,32,35-50Hzで基線と区別がつかなくなった。
考察
マダイの10℃のERG臨界融合頻度は26Hzに存在すると考えられる。20℃のERG臨界融合頻度は26Hzと判定された。25℃のERG臨界融合頻度は35Hzに存在すると考えられる。水温の上昇に伴う臨界融合頻度の増加が見られ,水温の高い方が運動視覚に優れると考えられる。
第2節 ストロボ光によるコイのERG臨界融合頻度
緒言
前節までマダイを用いてERGを記録した。本節では実験魚としてコイを用い,運動視覚の指標としてERG臨界融合頻度をマダイと比較できるか否かを検討する。
方法
実験は平成10年7月23日から12月31日にかけて行った。実験装置および測定条件の設定は前節と同様である。実験魚としてコイ6尾を用いた。
結果
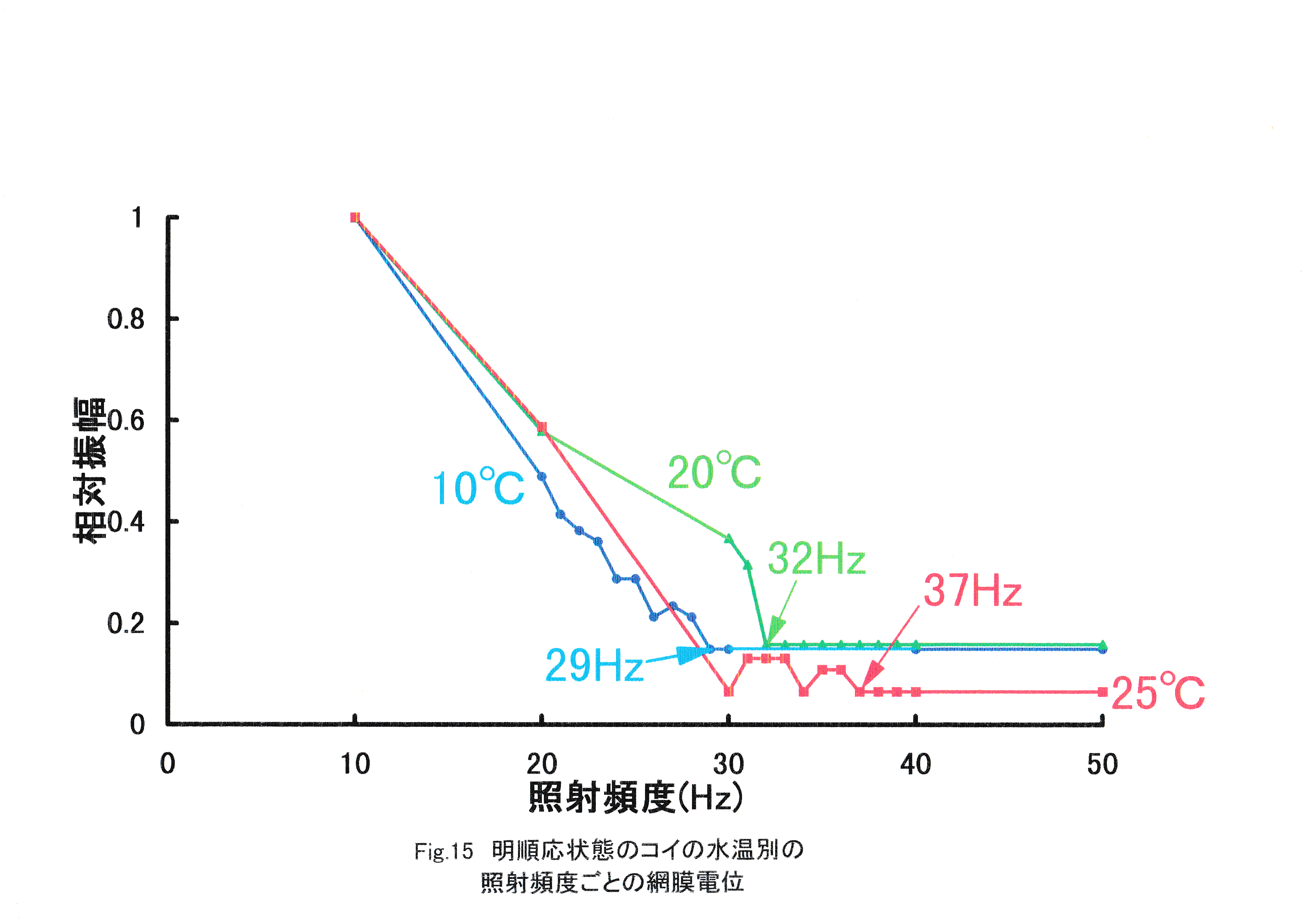
Fig.15に明順応状態のコイの水温別照射頻度ごとの網膜電位を示す。 全実験魚で照射頻度の増加に伴う網膜電位の振幅の減少が見られた。水温10℃の実験魚のERGは29-50Hzで基線と区別がつかなくなった。水温20℃の実験魚のERGは32-50Hzで基線と区別がつかなくなった。水温25℃の実験魚のERGは30,34,37-50Hzで基線と区別がつかなくなった。
考察
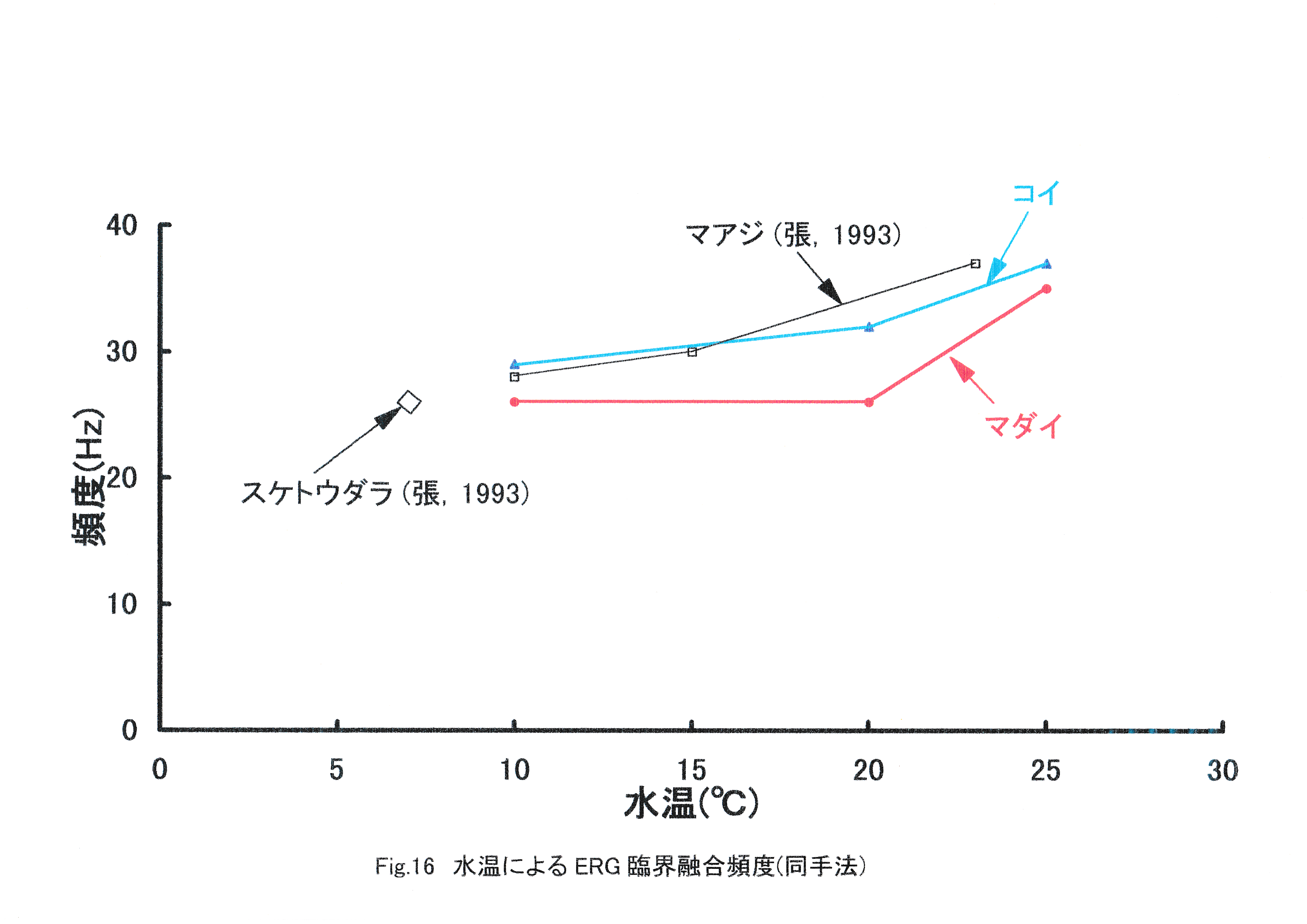
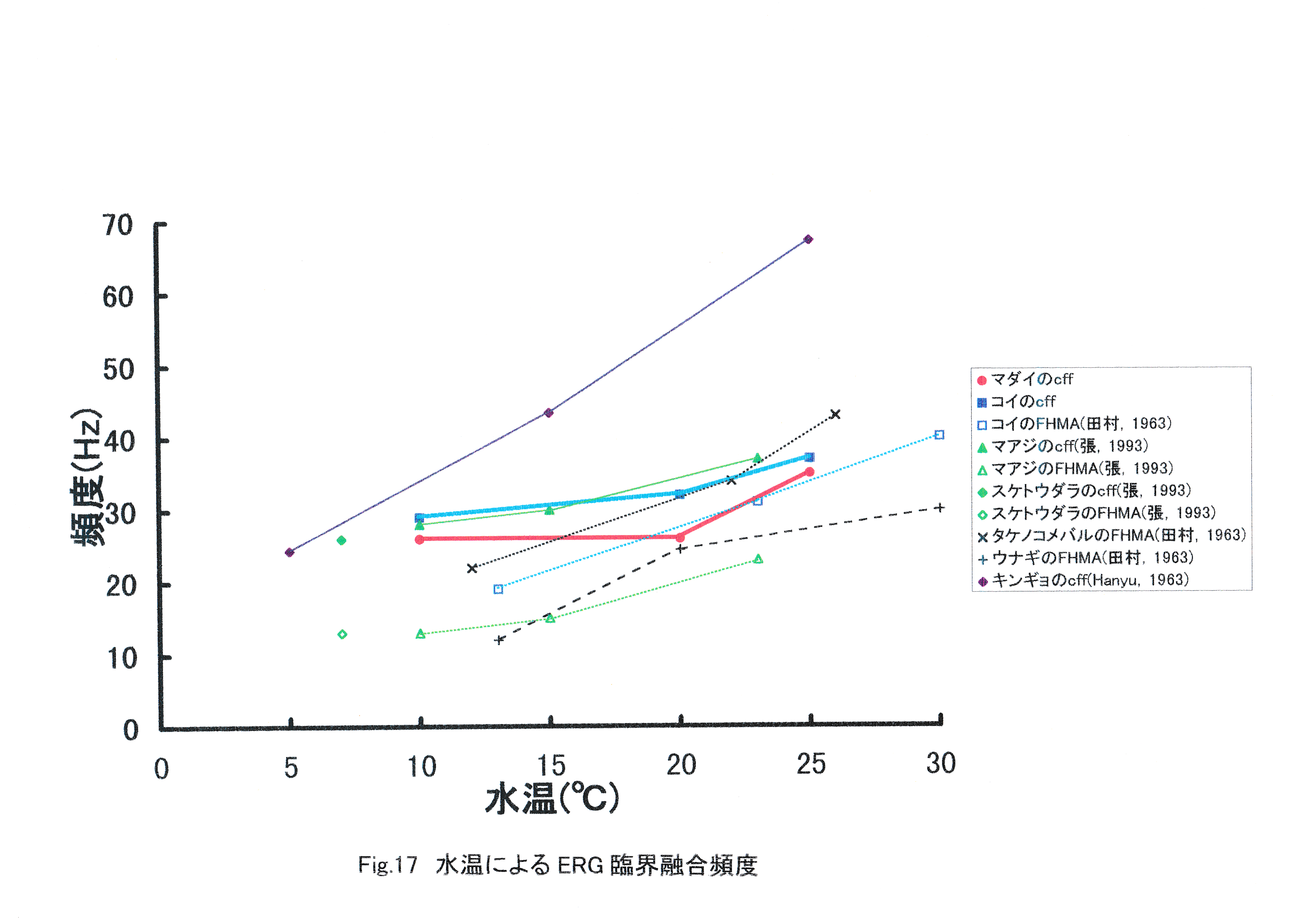
コイの臨界融合頻度は水温10℃で29Hz,水温20℃で32Hz,水温25℃で37Hzに存在すると考えられる。このようにコイでもマダイと同じく,水温の上昇に伴う臨界融合頻度の増加が見られたことから,魚類では水温の上昇に伴い運動視覚も向上すると考えられる。次に,本実験と同じ実験手法で得られたマアジとスケトウダラの水温ごとのERG臨界融合頻度を示し,マダイ,コイとの比較を行った(Fig.16)。 水温10~25℃においては,マダイよりもコイの臨界融合頻度の方が高くなった。一般にマダイ種苗は水温18.2~28℃において,コイは水温10~30℃において活発に採餌活動を行うとされている(山口,1978)。マダイよりもコイの臨界融合頻度の方が高い要因として,コイの適性水温がマダイより低温の広い範囲にわたっていることが挙げられる。マアジ,スケトウダラのERG臨界融合頻度はマダイ,コイと近似した値が得られ,同様に水温によるERG臨界融合頻度の向上という結果が示されている(張,1992)。さらにFig.17にその他の実験手法を含む魚類のERG臨界融合頻度を示す。 これまで測定された魚種としてタケノコメバル(田村,1963),ウナギ(田村,1963),キンギョ(Hanyu,1963)がある。これらは1960年代の実験であり,刺激として用いた光の条件,そして波形確認のための装置が異なり,本研究と比較が困難になっている。しかし,すべての報告において全体的に水温の上昇に伴う臨界融合頻度の増加が見られたことから,魚類では水温の上昇に伴い,運動視覚が向上すると考えられる。
総合考察
光強度の増加に伴う網膜電位の振幅の増大が見られた。ただし,照射開始直後の網膜電位の振幅は光強度の増加に伴って増大しなかった。照射開始直後の網膜電位の振幅が最も大きく,それ以降は一旦減少する傾向が見られた。それからさらに時間がたつにつれて,網膜電位の振幅は光強度の増加に伴って増大するようになった。最初の照射5回までで完全な暗順応状態となっていた網膜が光エネルギーを受けることで慣れたことによると考察した。これを確認するには組織生理学的手法を用いて,そのときどきの網膜の順応状態をもとに考察する必要があると考えられた。
また,明順応過程において網膜電位の振幅の増大が,暗順応過程において網膜電位の振幅の減少が見られた。本来ならば,明順応過程において網膜電位の振幅の減少が,暗順応過程において網膜電位の振幅の増大が見られるはずである。これは本実験の設定が明順応過程を知るには暗過ぎ(背景光が必要だった)て,暗順応過程を過程を知るには明るすぎた(照射する間隔を開けるべきだった)ことによると思われる。今後は明順応過程をたどるのに必要な背景光の光強度を知るための実験や,実験魚が暗順応過程をたどるのに必要な時間間隔を知る実験の必要性が示唆された。
暗順応状態より明順応状態で網膜電位の振幅は大きく,低頻度から高頻度という順序で照射した場合に照射頻度の影響を特徴的に示した。暗順応状態より明順応状態で網膜電位の振幅が大きくなったことから,ここでも組織生理学的手法を用いて,そのときどきの網膜の順応状態をもとに考察する必要があると考えられた。
電気生理学的実験で麻酔薬MS222を用いる場合は影響があるとされ,現在ではMS222は使用されなくなっている。本研究では全実験でMS222より影響が少ないとされる麻酔薬FA100を使用した。ただし,この薬の実験に与える影響は調べられてはおらず,今後調べる必要がある。
本実験で刺激光として用いたストロボスコープは閃光周波数1.4~440Hzの範囲でレンジの切り換えにより連続的に調整できるようになっている。ただし,照射頻度を上げると刺激光強度も増加してしまう。特に照射頻度を59Hzより高くして使用するときはレンジによっても刺激光強度が変わってしまうので注意が必要である。今後の実験では照射頻度の増加に伴う光強度の増加をどう押さえるかが問題であろう。
コイの臨界融合頻度は10℃で29Hz,20℃で32Hz,25℃で37Hzとなり,マダイの臨界融合頻度は10℃で26Hz,20℃で26Hz,25℃で35Hzとなった。ここで水温の上昇に伴う臨界融合頻度の増加が見られたのはなぜかという問題がある。水温が高くなれば視細胞の活動も活発になり,より高い時間的分解能を得られることによると考察したが,光照射によるERGの発生はけっきょくロドプシンなど感光色素の褪色分解に基づくものであり,この機構はまだ完全には判明していない。また最近は閃光刺激によって感光色素の褪色過程と密接な関係をもつ早期視細胞電位(early receptor potential, ERP)が発見され,コイやキンギョでも記録されている。今後はこれらの関係も含めて魚類のERG研究の展開が期待されるだろう(小林博,1970)。
謝辞
本研究のとりまとめに際して,平素から御指導御鞭撻を賜りました東京水産大学海洋生産学科魚群制御学研究室 有元 貴文 教授に有り難く厚くお礼申し上げます。
実験遂行において御助力,御教示を頂いた同研究室 秋山清二 助手ならびに 田原陽三 顧問に心より感謝申し上げます。
本論文の審査にあたり,副査をお引き受け頂きました同大学航法システム工学研究室 中村善彦 教授,同大学機能材料設計学研究室 稲田博史 助教授に深謝いたします。
そして,魚群行動学研究室院生 角田篤弘 氏,塩原 泰 氏を始め,同研究室の諸兄姉に感謝の意を表します。
参考文献
Ali,M.A. and Kobayashi, H.(1967)
Temperature : influence on the electroretinogram flicker fusion frequency of the sunfish(Lepomis gibbosus L.).
Rev.Can.Biol.,26:341-345.
Ali,M.A. and Kobayashi, H.(1968)
Electroretinogram flicker fusion frequency in albino trout.
Experientia,24:454-455.
有元貴文・安 永一(1998)
光による行動制御の技術
沿岸の環境圏,平野敏行監修,フジ・テクノシステム,pp.443-449
Norman,R.A. and Werblin,F.S.(1974)
Control of retinal senstivity.1.light and dark adaptation of vertebrate rods and cones.
J.Gen.Physiol.63:37-61.
安永一(1993)
威嚇光刺激に対する魚類の反応とその制御に関する基礎的研究
東京水産大学博士学位論文, pp.113-135.
安永一・有元貴文(1994)
ストロボ光に対するマアジの回避行動
Nippon Suisan Gakkaishi 60(6), pp.713-718.
Gruber, S. H.(1975)
Duplex vision in elasmobranchs : histological , electrophysiological and psychophysical evidence. in “Vision in Fishes”, New Approaches in Research(ed. M.A.Ali),Plenum Press, New York, London, pp.525-540.
羽生 功.(1972)
火光利用漁業の光源とその操法について.
日本水産学会誌,38:919-920
Hanyu, I. and Ali M. A. (1963)
Flicker Fusion Frequency of Electroretinogram in Light-Adapted Goldfish at Various Temperatures
SCIENCE, VOL.140, pp.662-663.
Hanyu, I. and Ali M. A. (1964)
Electroretinogram and its flicker fusion frequency at different temperatures in light adapted salmon(Salmo salar).
J. Cell Comp.physiol.63:309-321.
川村軍蔵・下和田隆(1983)
イシダイの帯模様弁別能
Bulletin of the Japanese Society of Scientific Fisheries, 49(1), 55-60
神作(1969)
CFF.感覚・知覚心理学ハンドブック.(和田陽平ら編)
東京,誠信書房.pp.238-249.
北村章二(1987)
カサゴのERG(網膜電図)-Ⅰ暗順応時における刺激と反応の関係.
Bull. Natl. Res. Inst. Aquaculture, 12, 53-60 .
北村章二(1988)
カサゴのERG(網膜電図)-Ⅱ明順応時による反応特性の変化.
Bull. Natl. Res. Inst. Aquaculture, 14, 139-145.
小林 博(1970)
§5. ちらつき光とERG,21.ERGと魚眼の機能
魚類生理;川本信之編;恒星社厚生閣,pp.452-461.
小林 博(1963)
メクラウナギの痕跡眼に見られる活動電位,
動物学雑誌,72:6-12
Kobayashi, H. (1962)
A comparative study on electroretinogram in fish, with special reference to ecological aspects.
J. Shimonoseki Coll. Fish., 11:17-148.
丘 直通(1935)
動物の最小識時に就て
科学,第5巻,第2号
DAVID R.SAGER and CHARLES H.HOCUTT(1987)
Estuarine Fish Responses to Strobe Light,Bubble Curtains and Strobe Light/Bubble-Curtain Combinations as Influenced by Water Flow Rate and Flash Frequencies
Fisheries Research,5 pp.383-399.
Schmidt, R. F.(1989)
Fundamentals of sensory physiology. (ed. By R. F. Schmidt), (transl. by Y. Iwamura et al.),
Kinpodo, Kyoto, pp.165-166.
宗宮弘明・丹羽 宏(1991)
視覚.
魚類生理学. (板沢靖男, 羽生 功編). 恒星社厚生閣, pp.403-441.
田村 保(1970)
視覚.
魚類生理. (川本信之編), 恒星社厚生閣, pp.423-451.
田村 保(1963)
魚の眼の機能の研究方法
Bulletin of the Japanese Society of Scientific Fisheries,
Vol.29,No.1.
田村 保(1957)
魚類の視覚について,Ⅱ.視認識,2. 運動視覚
水産学集成,東大出版会.
Tamotsu Tamura and Isao Hanyu(1959)
THE FLICKER ELECTRORETINOGRAM OF THE CARP EYE
Bulletin of the Japanese Society of Scientific Fisheries, Vol.25, No.10-12, pp.624-631.
TSUNEO TOMITA and TAKASHI YANAGIDA(1981)
ORIGINS OF THE ERG WAVES
Vision Research Vol.21.pp.1703-1707.
張秀梅(1992)
魚類の視覚生理とトロール漁法への応用に関する研究
東京水産大学博士学位論文, pp.85-220.
張秀梅(1995)
トロール漁法の漁獲過程
水産学シリーズ108,日本水産学会監修,魚の行動生理学と漁法,
有元貴文・難波憲二編,恒星社厚生閣刊,pp.21-30.
張秀梅・有元貴文(1993)
ストロボ光によるマアジのERG臨界融合頻度
Journal of Tokyo University of Fisheries, Vol.80, No.1, pp.61-67.
張 秀梅・有元貴文・井上 実(1989)
ストロボ光によるマアジの網膜運動反応
Journal of the Tokyo University of Fisheries, Vol.76, No.1-2, p.65-72.
山口正男(1978)
養殖場の適地条件
タイ養殖の基礎と実際,恒星社厚生閣
米沢勝之・田村 保(1971)
チラツキ刺激に対する魚類網膜活動電位の研究
Bulletin of the Japanese Society of Scientific Fisheries, Vol.37, No.9, pp.846-850.
Views: 192


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}